Условия, при которых стимулируются слуховые нервные волокна.
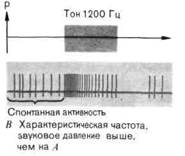
Каждое волокно в слуховом нерве начинается от узкой ограниченной области улитки, в некоторых случаях от одной внутренней волосковой клетки. Поскольку отдельные участки улитки связаны с отдельными частотами, каждое нервное волокно оптимально возбуждается звуком определённой_.частоты. Эта частота называется характеристической частотой данного волокна. Таким образом, волокно в слуховом нерве подвергается возбуждению легче всего, когда ухо стимулируется звуком частоты, характеристичной для данного волокна (рис. 5-14). Чистые тоны очень низкой интенсивности возбуждают специфические одиночные волокна нерва. Если же ухо стимулируется тонами, частоты которых не являются характеристическими, то оно может быть активировано только при достаточном увеличении интенсивности. Это свойство «настройки» показано на рис. 5-15, где на графике для каждой частоты дано звуковое давление, которое вызывает околопороговый ответ в каждом из двух волокон, различающихся по характеристической частоте (~ 3000 и 4500 Гц). Затененный участок над каждой кривой показывает диапазон частот и интенсивностей, в котором волокно реагирует. Такие кривые называются кривыми настройки. Если звуковой стимул содержит несколько частот, то активируются все соответствующие группы нервных волокон. Длительность стимула кодируется длительностью активации, а интенсивность - степенью активации. При повышении уровня звукового давления не только усиливается возбуждение данных волокон (увеличивается частота их разряда), но в активность вовлекаются также дополнительные волокна (с частотами, близкими к характеристической). На рис. 5-15 показано также, что в ответ на звуковой стимул, параметры которого лежат в области перекрывания, возбуждаются оба волокна.
Итак, можно сказать, что на уровне первичных афферентных волокон звуковой стимул разлагается на свои частотные компоненты. На более высоких уровнях слухового пути переработка информации носит совсем иной характер.
Анатомия слухового пути.На рис. 5-16 приведена сильно упрощенная схема основных частей слухового пути. Для большей ясности показаны пути только от левого уха. Кончиками стрелок обозначены точки переключения, где нейроны образуют синапсы с нейронами более высокого порядка; на схеме не различаются тормозные и возбудительные синапсы. Первичные афферентные волокна идут сначала к кохлеарному ядру, подразделенному на вентральную и дорсальную части. От вентральной части вентральный тракт идет к ипси- и контралатеральным оливарным комплексам. Таким образом нервные клетки в каждом оли-варном комплексе получают входы от обоих ушей. Здесь расположен самый нижний из тех уровней головного мозга, на которых возможно сравнение акустических входов от обоих ушей. Такого рода сравнение производится в особенности в добавочном ядре-в центре, который уже был упомянут в разд. 5.2 в связи с ощущением направленности звука. Дорсальное кохлеарное ядро служит началом дорсального тракта. Эти волокна переходят на противоположную сторону и там образуют синапсы с нейронами ядра латерального лемниска. Одни восходящие волокна клеток оливарного комплекса проецируются на ту же сторону, другие образуют перекрест. Над латеральным лемниском слуховой тракт идет через два ядра переключения - нижний бугорок и медиальное коленчатое тело~к своему последнему назначению-первичном слуховой коре в височной доле. Таким образом, этот тракт состоит из последовательности по меньшей мере пяти или шести нейронов. В действительности в нем имеются еще и другие переключения, не показанные на данной схеме, так что возможны еще более длинные цепочки. Кроме того, слуховой путь отдает много коллатералей. Наконец, слуховая система содержит не только афферентные, центростремительные волокна, но также эфферентную, центробежную систему, не показанную на рис. 5-16; функция этой эфферентной системы не совсем ясна.
Характеристики ответов центральных слуховых нейронов.Нейроны слухового нерва возбуждаются совсем простыми стимулами, например чистыми тонами, но для нейронов высших уровней слухового пути обычно требуются более сложные звуки. Хотя в вентральном кохлеарном ядре клетки ведут себя так же, как и нейроны слухового нерва, в дорсальном кохлеарном ядре их ответы заметно отличаются. В определенных условиях происходит торможение этих нейронов входными слуховыми сигналами. Торможение создается обширными реципрокными связями между нейронами.
При отведении от клеток еще более высоких уровней ответы на чистые тоны часто вообще отсутствуют. Эти клетки отвечают на сложные звуковые паттерны, например на амплитудно- или частотно-модулированные звуки, т.е. звуки с непрерывно меняющейся интенсивностью или частотой. Другие нейроны реагируют только на начало или только на конец звукового стимула и т.п. Часто клетка активируется одними частотами и тормозится другими. B общем можно сказать, что чем выше уровень, на котором находится нейрон в слуховом пути, тем более сложные звуки вызывают его возбуждение. Раньше думали, что частотная избирательность нейронов, отвечающих на чистые тоны, усиливается на высших уровнях, но в действительности это не так. На высших уровнях настройка нейронов не намного острее, чем на уровне слухового нерва; собственно говоря, многие клетки в слуховой коре могут быть активированы очень широкой полосой частох_В то же время в некоторых участках слуховой коры имеется тонотопическая карта улитки.

|

|
|
Рис. 5-14. Разряд в одиночном волокне слухового нерва в ответ на стимуляцию его характеристической и близкой частотами при двух разных интенсивностях.

 Рис. 5-15. Кривые настройки двух волокон слухового нерва. См. текст
Рис. 5-15. Кривые настройки двух волокон слухового нерва. См. текст

 Рис. 5-16. Сильно упрощенная схема слухового пути, описанного в тексте.
Рис. 5-16. Сильно упрощенная схема слухового пути, описанного в тексте.
В обычной жизни звуки редко бывают чистыми тонами. Содержат разнообразные компоненты подобно сложным звукам, упомянутым выше. Это особенно верно для речи, структура которой все время меняется; некоторые ее элементы очень короткие, и, как правило, в ней возникает одновременная модуляция амплитуды и частоты. Очевидно, нейроны в высших отделах слухового пути специализированы для извлечения специфических характеристик таких сложных звуков, например изменения частоты в произносимом слове, и тем самым для узнавания некоторых констелляций стимулов. Клетки, отвечающие на такие особые констелляции, являются слуховыми аналогами сложных или сверхсложных нейронов зрительной коры (см. разд. 4.3). В обеих модальностях такие клетки извлекают определенные свойства стимула, участвуя в процессе опознания его структуры.
Другой путь исследования нейронной переработки звукового сигнала состоит в регистрации потенциалов электродами, фиксированными на коже головы. Эти потенциалы представляют собой суммарную активность многих нейронов, но они сильно ослаблены и для их анализа нужны ЭВМ. Такие измерения образуют основу так называемой аудиометрии по вызванным потенциалам (АВП); этот метод недавно стали применять как способ диагностики локализации поражения в случаях потери слуха находится ли оно на уровне улитки или выше.
Адаптация в слуховой системе.Явление адаптации присуще всем сенсорным системам, и слуховая система не составляет исключения. Это свойство, как периферических рецепторов, так и центральных нейронов. Адаптация к данному уровню звука приводит к повышению слухового порога, но из этого не следует, что она нежелательна или бесполезна. Адаптация к некоторым интенсивностям и частотам может в действительности понизить дифференциальный порог, усилив различение разных уровней слухового давления. Так, например, тон в 4000 Гц на рис. 5-6 при 33 дБ УЗД имеет уровень громкости 40 фонов, а тот же тон при 52 дБ УЗД-60 фонов. Следовательно, в этой части графика, чтобы получить разницу уровня громкости в 20 фонов, нужно усиление звукового давления на 19 дБ. Но если ухо адаптировано к долгому воздействию 4000 Гц, то изофоны в этой области смещаются вверх; в частности, для низких уровней громкости изофоны сместятся дальше, чем для высоких. В результате разница в уровне громкости в 20 фонов потребует изменения меньше чем на 19 дБ УЗД, например на 14 дБ. Иными словами, в адаптированном состоянии данная разница в уровне звукового давления вызовет большую разницу в уровне громкости. Субъективная разница становится больше, и, следовательно, порог различения понижается.
Предыдущее изложение могло создать впечатление, что ощущение тона основано полностью на локализации смещения в улитке, зависимой от частоты, как это постулирует теория места, или на настройке волокон слухового нерва на их характеристические частоты. Однако не все психофизические наблюдения над тональными ощущениями можно объяснить одной только теорией места. Так, например, при стимуляции уха последовательностью коротких импульсов давления (щелчков) с интервалами в 5 мс, как и следовало ожидать, слышится тон в 200 Гц. Но если предварительно профильтровать звук так, чтобы пропускались частоты только выше определенного уровня (скажем, 600 Гц), то в стимуле уже не будет частоты 200 Гц. Однако субъективно воспринимаемый тон по-прежнему будет 200 Гц. Это явление еще не получило убедительного объяснения, но его нельзя объяснить одной только теорией места. По-видимому, мозг «воссоздает» недостающую основную частоту информации, которая содержится в сохранившихся обертонах.
В 5.14. Предположим, что звук активирует волокна слухового нерва, обладающие несколькими разными характеристическими частотами - около 800, 1600 и 3200 Гц. Волокна с характеристическими частотами, лежащими между этими величинами, не активируются. Причиной этого может быть
а) то, что звук содержит несколько частот;
б) то, что интенсивность звука очень велика;
в) то, что нейроны слухового нерва обладают спонтанной
активностью.
В 5.15. Слуховой путь содержит среди других следующие ядра переключения.
а) Кохлеарное ядро.
б) Ретикулярная формация.
в) Верхний бугорок.
г) Медиальное коленчатое тело.
д) Латеральное коленчатое тело.
е) Слуховая кора.
ж) Верхняя олива.
з) Чечевицеобразное ядро.
В 5.16. Возбуждение первичных афферентных волокон в слуховом нерве может быть вызвано
а) любым звуковым стимулом, лишь бы он был выше порога;
б) только чистыми тонами;
в) только сложными звуками, например частотно- или ампли-
тудно-модулированными тонами;
г) только началом или концом звука («on»- и «оff»-нейроны).
6. ФИЗИОЛОГИЯ ЧУВСТВА РАВНОВЕСИЯ
Дата добавления: 2015-10-05; просмотров: 1472;