Часть III ЭВОЛЮЦИЯ ПСИХИКИ. 6 страница
В первой и второй частях этой книги рассматривалось преимущественно поведение высших позвоночных. По этой причине мы ограничимся здесь лишь некоторыми дополнениями к сказанному.
Локомопия.
Мышечная система позвоночных состоит из симметрично расположенных вдоль тела сегментов. Правда, посегментное расположение мышц выражено у хордовых слабее, чем у членистоногих и червей, а у высших представителей позвоночных оно сильно нарушено. Различают соматическую мускулатуру, обслуживающую эффекторные органы, и висцеральную — мускулатуру внутренних органов и кожи. Соматическая мускулатура состоит всегда из поперечнополосатых мышечных волокон.
Конечности позвоночных представлены непарными и парными образованиями. Непарные конечности имеются только у круглоротых и рыб. Это спинной, заднепроходный и хвостовой плавники. Парные конечности, передние и задние, сильно различаются у разных позвоночных соответственно своей функции (плавники, крылья, лапы, ласты, ноги, руки), однако сравнительное изучение строения их скелета позволяет отчетливо проследить эволюционные преобразования от общей примитивной начальной формы.
1 Северное А.Н. Главные направления эволюционного процесса. Морфобиологическая теория эволюции. 3-е изд. М., 1967. С. 115.
Основная функция конечностей состоит у всех животных в локомоции, в перемещении животного в пространстве. Однако у многих членистоногих и позвоночных сюда добавляется еще функция опоры приподнятого над субстратом тела. Поэтому в этих случаях говорят об опорно-локомоторной функции конечностей. Не вдаваясь здесь в детальное рассмотрение разнообразных форм этой основной функции конечностей позвоночных, укажем лишь на некоторые существенные моменты.
Видный советский ученый Н.А.Бернштейн писал, что постепенно назревшая в филогенезе потребность в быстрых и мощных движениях — привела на одной из его ступеней к возникновению и параллельному развитию «костно-суставных кинематических цепей скелета» и поперечнополосатой мускулатуры, снабженной соответственными нервными образованиями. У позвоночных эти скелетно-мышечные системы («неокинетические системы», по Бернштейну) получают существенное прогрессивное развитие по сравнению с членистоногими, причем особенно это относится к рассматриваемым здесь высшим позвоночным, в частности к их локомоторным способностям. Бернштейн указывает в этом отношении на глубокие качественные различия между низшими и высшими позвоночными, обусловленные усложнением возникающих перед организмом двигательных задач, возрастанием разнообразия реакций, требующихся от организма, более высокими требованиями в отношении дифференцирован-ности и точности движений. «Достаточно напомнить, -пишет Бернштейн, — насколько, например, аэродинамический полет птицы сложнее почти полностью гидростатического плавания рыбы или насколько богаче по контингентам участвующих движений охота хищного млекопитающего по сравнению с охотой акулы. Молодая отрасль проворных теплокровных млекопитающих победила тугоподвижных юрских завров именно своей более совершенной моторикой»2.
2 Бернштейн Н.А. О построении движений. М., 1947. С. 9.
В современных этологических исследованиях локомоторная активность изучается в ее видотипичных проявлениях как адаптация к специфическим условиям существования: разновидности и особенности ходьбы, бега, прыганья, лазанья, плавания, полета и т.д. определяются особенностями образа жизни и служат важным приспособлением к окружающей среде. Вместе с тем всем формам локомоции присуща ритмичность, выражающаяся в том, что движения выполняются в четкой последовательности многократно и относительно стереотипным образом (хотя поведение животного в целом при этом не является стереотипным). Эта ритмичность основана на эндогенной центрально-нервной стимуляции и проприоцепторной обратной связи. Помимо проприоцепторной чувствительности внешние импульсы лишь регулируют эти ритмы, соотносят их параметры (сила, скорость, длительность движений и т.д.) с конкретными условиями тех ситуаций, в которых оказывается животное. В частности, внешние раздражения вызывают начало или конец локомоторных движений, хотя и это может произойти в результате эндогенной стимуляции.
Сказанное достаточно поясняет тот факт, что локомоторные движения относятся к наиболее «автоматизированным» и однообразно выполняемым компонентам всей двигательной сферы животных. В этой же связи стоит и относительная малочисленность форм локомоции у каждого вида. Определяющей для локомоции является ее физическая, механическая функция. Сами локомоторные движения дают животному только минимальную информацию об окружающем мире.
При этом необходимо, правда, учесть, что локомоторная активность включает в себя и ориентировочные компоненты, имеющие, конечно, и определенное познавательное значение. Так, например, прыгающие животные, особенно древесные, должны перед прыжком точно «рассчитать» расстояние. Как показали советские исследователи поведения животных В.М.Смирин и О.Ю.Орлов, это делается с помощью особых движений «взятия параллакса» (рис. 42). Оказавшись в новом месте, летяга «прицеливается» к разным предметам, это же она делает перед каждым прыжком, хотя со временем число таких движений уменьшается. В итоге уходящее от опасности животное придерживается заранее «отработанного» пути без лишних движений и совершает прыжки с поразительной точностью.

|
Рис. 42. Специфические движения летяги для определения удаленности предметов. Летяга «прицеливается» перед прыжком: приподнявшись, животное производит боковые движения головой в горизонтальной плоскости. Подобные ориентирующие движения животные производят особенно часто при передвижении в незнакомой местности (по Смирину и Орлову).
Манипулирование.
Эффекторные органы позвоночных всегда выполняют сразу несколько функций — помимо основной функции еще большее или меньшее число дополнительных. В этом проявляется мультифункциональность этих органов. Что касается конечностей высших позвоночных, то как уже отмечалось, их основой является опорно-локомоторная функция: все же дополнительные их функции сводятся к разнообразным формам манипулирования предметами (пищевыми или несъедобными). Особый интерес представляют для зоопсихолога формы манипулирования передними конечностями, которые в процессе эволюции привели к орудийной деятельности приматов и тем самым стали важнейшей биологической предпосылкой зарождения трудовых действий у древнейших людей.
Говоря о манипуляциях, дополнительных функциях передних конечностей у высших позвоночных, необходимо иметь в виду, что сложные и разнообразные дополнительные функции присущи у этих животных и ротовому аппарату, причем существует далеко идущее функциональное взаимодействие между этими двумя основными эффек-торными системами. Поэтому целесообразно анализировать дополнительные функции передних конечностей и ротового аппарата в комплексе. Так, например, у взрослых лисиц обнаруживается 45 дополнительных функций передних конечностей и ротового аппарата, у барсука — 50, у енота — около 80, а у низших обезьян (макаков и павианов) — свыше 150 (данные Фабри).
Важно отметить, что по мере специализации млекопитающих в сфере локомоции часть дополнительных функций передних конечностей передается челюстному аппарату. Так, например, в ряду медведь — енот — барсук — лисица конечности лисицы в наибольшей степени приспособлены к продолжительному быстрому бегу. Но при этом дополнительные (манипуляторные) функции конечностей сводятся к минимуму и соответствующие жизненно необходимые движения (например, в сфере питания) выполняются почти исключительно ротовым аппаратом. Как раз наоборот обстоит дело у медведей.
Если иметь в виду познавательное значение манипулирования, то следует особенно выделить те его формы, при которых животное удерживает предмет перед собой свободно на весу обеими или одной конечностью. Такая фиксация предмета дает животному наилучшие возможности для комплексного (оптического, ольфакторного, тактильного, кинестетического и пр.) обследования, особенно в ходе деструктивного воздействия на него (практического анализа). По данным, полученным Фабри, это дает при известных условиях обезьянам, а отчасти и другим способным на это млекопитающим наиболее всестороннюю и разнообразную информацию, необходимую для развития высших форм психической активности. Оказалось, что медведи владеют тремя способами фиксации объекта на весу, еноты — шестью, низшие обезьяны и полуобезьяны — тремя десятками таких способов! К тому же только обезьяны обладают достаточно различными двигательными возможностями, чтобы произвести подлинный деструктивный анализ (расчленение) объекта на весу.
О формировании манипуляционной активности уже говорилось раньше (см. ч. II). Напомним лишь, что первым начинает функционировать челюстной аппарат, позже появляются движения передних конечностей, которые, однако, вначале еще нуждаются в поддержке более сильной системы челюстного аппарата. Впоследствии движения конечностей становятся все более сильными и самостоятельными и одновременно все более разнообразными, т.е. возрастает их мультифунк-циональность. Ряд функций, в частности хватательные, первоначально выполнявшиеся челюстями, переходит при этом к конечностям. Вершиной этого развития являются дифференцированные движения, выполняемые только одной конечностью. Как уже отмечалось, прогрессивное развитие дополнительных двигательных функций передних конечностей являлось и является важнейшим фактором психического развития.
Комфортное поведение. Сон и покой.
Разновидностью манипулирования можно отчасти считать комфортное поведение, служащее уходу за телом животного, с той лишь особенностью, что объектом манипулирования является не посторонний предмет, а именно собственное тело. Но, кроме того, сюда относятся и нелокализованные движения, производимые всем телом и лишенные специальной пространственной направленности. В итоге можно выделить следующие категории комфортных движений: очищение тела, потряхивание, почесывание (определенного участка тела об субстрат), катание по субстрату, купание (в воде, песке и т.д.).
Комфортное поведение широко распространено и среди членистоногих (достаточно вспомнить, как муха чистит лапками голову, крылья и т.п.), иногда встречаются и специальные морфологические образования или даже органы для очищения тела. Инстинктивная природа комфортных движений обнаруживается в видоспецифичности их выполнения, равно как в строгой фиксированности участков тела, очищаемых тем или иным органом и способом. Столь же четко видотипичными являются и позы сна и покоя. Так, например, у зубров, бизонов и их гибридов было описано всего 107 видотипичных поз и телодвижений по 8 сферам поведения (исследования М.А.Дерягиной). Наибольшее число поз (2/3) приходится на сферы сна, покоя и комфорта. При этом любопытно, что у детенышей (до 2—3 месяцев) всех трех изученных групп животных видотипичные различия в этих сферах еще не выражены. Напрашивается вывод, что эти различия формируются постепенно в более позднем возрасте.
Если рассмотреть одну из форм комфортного поведения, облизывание, то, согласно Дерягиной, оказывается, что только бизон облизывает стоя корень хвоста или приподнятую переднюю ногу, но только зубр облизывает стоя бедра задней ноги и т.д. Если не считать гибридов, то существует несколько способов облизывания, из них присущих только зубру — один, только бизону — четыре, общих зубру и бизону — четыре. Здесь опять проявляются четкие различия в инстинктивных движениях у близкородственных видов (зубр и бизон рассматриваются иногда даже как подвиды).
Сенсорные особенности.
В полом соответствии с высокоразвитыми эффекторными системами высших позвоночных находится и высокий уровень их сенсорных способностей. Наибольшее значение имеют органы слуха и равновесия (внутреннее ухо, начиная с земноводных — дополнительно среднее ухо, а у млекопитающих также и наружное ухо), обоняния (обонятельные мешки и раковины) и зрения (глазные яблоки). Хорошо развиты также кожная и мышечная (тактильно-кинестетическая) и термическая чувствительность, вкус, а в ряде случаев и другие виды чувствительности (электрическая, вибрационная и др.). Соответственно развиты у позвоночных и разные таксисы.
Сопоставление роли зрения и обоняния в жизни высших позвоночных показывает, что у большинства млекопитающих ведущую роль играет обоняние, которое достигает у них удивительной остроты (только акулы могут в этом отношении сравниваться с ними). Так, крот чувствует добычу сквозь толщу земли, белый медведь -сквозь лед. Бурый медведь чувствует на расстоянии 20 метров мед, зарытый в землю на глубину 50 сантиметров. Хорошо известны поразительные обонятельные способности собак и копытных.
Однако, как уже отмечалось, в каждом конкретном случае степень развития обоняния (и соответствующих морфологических структур в рецепторе и головном мозге) определяется особенностями биологии вида. Так, например, по Б.С.Матвееву, даже у родственных видов, обитающих в сходных условиях, но различающихся по биологии, — у обыкновенной полевки (Mictorus arvalis) и полевой мыши (Apodemus agrarius) - - наблюдаются существенные сенсорные различия. Полевые мыши поедают разнообразный корм - много семян, а также животный корм, полевки же - - преимущественно зеленый корм, который всегда имеется в избытке. Вследствие этого обоняние развито у полевой мыши сильнее, чем у полевки.
Зрение лучше всего развито у птиц и приматов. Оно играет важную роль в их пищедобывательной, оборонительной, воспроизводительной и других формах поведения, обеспечивая четкую ориентацию животного в пространстве. Особое значение приобретает зрение при дальней ориентации мигрирующих птиц, а также внут-ритерриториальной ориентации млекопитающих. Хорошо изучена, например, ориентация птиц по топографическим признакам (например, береговой линии), поляризованному освещению небосвода и астрономическим ориентирам - солнцу, звездам. Последний случай представляет собой наглядный пример ориентации на основе менотаксисов.
У многих млекопитающих ориентация на своей территории (индивидуальном участке или территории стада) осуществляется с помощью фото- и хемомнемотаксисов (по оптическим и ольфакторным меткам). Кроме того, даже у хуже видящих млекопитающих (например, куньих) зрение играет немаловажную роль при ориентации по подвижным объектам, особенно во время ловли добычи. Даже животные с определенно плохим зрением, как, например, моржи, которые над водой близоруки, пытаются ориентироваться с помощью не только обоняния, но и зрения, особенно опять-таки когда имеют дело с подвижными объектами (приближающимися к ним животными или людьми).
Наиболее мощным зрением, очевидно, во всем мире животных обладают хищные птицы. Достаточно сказать, что сокол способен при благоприятных условиях увидеть сидящего голубя на расстоянии почти полутора километров. Равным образом грифы находят свой корм (трупы животных) на огромном удалении с помощью зрения. Но они не в состоянии обнаружить прикрытые останки животных. Но это относится лишь к птицам, обитающим в степных просторах Восточного полушария. Грифы же Центральной и Южной Америки (например, черные американские грифы), разыскивающие пищу в густых лесах, не могут пользоваться зрением и находят ее, как это ни кажется невероятным, не менее успешно и с не меньшего расстояния с помощью обоняния. Аналогично обстоит дело и у млекопитающих. У копытных, обитающих на открытых пространствах, сильнее развито зрение, чем у их родственников, живущих в лесах, даже если животные принадлежат к одному и тому же роду (например, бизон -преимущественно степное животное и зубр — обитатель лесов).
Все эти примеры наглядно показывают, что образ жизни, биология всецело определяют характер поведения и психики животных (в отличие от человека, который благодаря своей общественно-трудовой деятельности достиг далеко идущей независимости от биологических1 факторов и психическая деятельность которого обусловливается социальными условиями его жизни). В приведенных примеpax это положение материалистической зоопсихологии относится конкретно к сенсорной сфере животных, в частности к вопросу о ведущей рецепции. Как мы видели, в каждом отдельном случае, даже у близкородственных видов, она определяется конкретными условиями жизни животного. Но то же самое относится и к психическому отражению в целом, характер которого также всегда определяется этими условиями.
Зрительные обобщения и представления.
Подлинная рецепция, истинное восприятие предметных компонентов среды как таковых возможны лишь на основе достаточно развитой способности к анализу и обобщению, ибо только это позволяет полноценно узнавать постоянно меняющие свой вид (и другие свойства) предметные компоненты среды.
Исследования, проведенные на разных видах позвоночных (помимо круглоротых), показали, что все они способны к предметному восприятию, в частности к восприятию форм. Уже на этом основании можно заключить, что все позвоночные находятся на стадии перцептивной психики. Однако внутри этой стадии наблюдаются существенные различия между низшими и высшими позвоночными.
Так, рыбы способны в эксперименте отличать геометрические фигуры, например квадрат от треугольника, независимо от меняющейся величины этих фигур. Но достаточно в контрольном опыте перевернуть треугольник (острием вниз) или квадрат (придать ему положение ромба), как рыба перестает узнавать эти фигуры. Следовательно, восприятие и узнавание формы оказываются здесь ограниченными, недостаточно гибкими. Отсутствует способность к быстрому переносу сформировавшегося зрительного представления на основе обобщения. Млекопитающие же вполне способны к такому обобщению и легко узнают треугольник или другую геометрическую фигуру любой величины в любом положении. Существенно превосходят рыб также птицы. Правда, и курица, по некоторым данным, не узнает перевернутый треугольник, которы до этого безошибочно отличала от другой геометрической фигуры. Вместе с тем курица способна к значительным обобщениям, опознавая, например, геометрические фигуры независимо от их цвета. Более того, будучи обучена выбирать сплошной треугольник, она также реагирует на него, когда он предъявляется ей в виде штриховки или даже лишь намечен тремя точками! Так же как и рыбы, птицы способны распознавать относительную величину фигур и так же ошибаются на основе зрительных иллюзий. Способность к зрительному обобщению у многих позвоночных экспериментально доказана. Особенно в отношении внешних их представителей было показано, что они в состоянии даже в очень сложных ситуациях вычленять существенные детали в воспринимаемых объектах и узнавать эти объекты в сильно измененном виде. Отсюда напрашивается вывод о существовании у позвоночных достаточно сложных общих представлений. Представления чрезвычайно важны для выживания. В них закрепляется в общем виде индивидуальный опыт, что позволяет животному легче ориентироваться в существенно меняющейся окружающей среде при временном отсутствии жизненно важных раздражителей. Подобные зрительные обобщения обнаруживаются, в частности, в явлениях «экстраполяции», описанных Л.В.Крушинским, которые проявляются в том, что в ряде случаев животные способны, проследив за движением какого-либо предмета, ориентироваться затем на ожидаемый путь его перемещения. Следовательно, учитывается предстоящее передвижение объекта после его исчезновения. В проведенных им опытах птицы и млекопитающие помещались перед туннелем, имевшим посредине разрыв, через который можно было наблюдать за движением перемещающейся в туннеле кормушки с приманкой. В других опытах применялась ширма со щелью. Некоторые из подопытных животных (врановые, хищные) обегали затем туннель или ширму по направлению движения приманки и ожидали ее в месте появления. На такое «предвосхищающее» поведение оказались, однако, способными лишь животные, имеющие в естественных условиях дело с прячущейся добычей, которую приходится выслеживать или преследовать, зачастую обгоняя ее, или же сами прячущие корм про запас. Как уже раньше указывалось, мы имеем здесь дело с широко распространенной среди высших позвоночных способностью к нахождению обходного пути. При этом важно отметить, что в любом случае, когда животное огибает непрозрачную преграду или даже просто отворачивается от целевого объекта, оно на время теряет последний из виду. Более того, каждое представление по самой природе своей требует отсутствия ранее воспринятых объектов.
Убедительные доказательства тому, что обобщенные зрительные образы в форме представлений широко распространены среди позвоночных, дали эксперименты, проведенные методом отсроченных реакций (исследования В.С.Хантера, Р.Йеркса, О.Л.Тинклпоу, Ф.Бойтендайка, В.Фишеля и др.). В Советском Союзе такие эксперименты проводились Н.Ю.Войтонисом, И.С.Беритовым, Г.З.Ро-гинским и др. Обычная схема проведения подобных опытов такова: на виду у животного прячется корм, который, однако, позволяется ему отыскать лишь некоторое время спустя. Обычно подопытное животное должно произвести выбор между несколькими местами, где может находиться спрятанная приманка. При успешном решении задачи имеет место реакция на отсутствующий стимул, что возможно лишь при наличии чувственных представлений.
Опыты по методу отсроченных реакций с успехом ставились над различными позвоночными, особенно же часто над обезьянами. Весьма эффектными были опыты, которые проводил Тинклпоу, когда на глазах у шимпанзе прятались фрукты (бананы), которые затем незаметно для животного подменялись значительно менее привлекательным кормом — салатом или капустой. Найдя спустя положенное время зелень, обезьяна медлила брать ее и продолжала, иногда с визгом, поиски. Последние продолжались до 33 секунд, в то время как в контрольном опыте, когда обезьяне с самого начала показывался салат, который затем не подменялся, поиски продолжались лишь 3 секунды и обезьяна спокойно съедала его. Напрашивается вывод, что обезьяна ищет виденный ею вначале объект, руководствуясь зрительным представлением, которое, по Тинклпоу, отражает также определенные количественные и качественные показатели.
Войтонис на основании своих экспериментов также пришел к выводу, что обезьяны (макаки и шимпанзе) реагируют не на кормушку, содержащую приманку, а на корм как таковой, точнее, на специфический вид корма. То же самое относится и к собакам, но не к рыбам (опыты Бойтендайка и Фишеля).
Наличие у позвоночных (а возможно, и у некоторых высших беспозвоночных) представлений, выражающихся в отсроченных реакциях и способности к нахождению обходных путей (включая и явления экстраполяции), придает их поведению исключительную гибкость и намного повышает эффективность их действий на поисковых этапах поведенческих актов. Вместе с тем не следует переоценивать эти способности, ибо они не обязательно свидетельствуют о высоком уровне психического развития. Основываясь на развитой памяти, они встречаются у позвоночных на разных филогенетических ступенях в зависимости от образа жизни животного: наряду с животными, прячущими в разных местах запасы, эти способности свойственны хищникам, охотящимся на труднодосягаемых, прячущихся или лишь эпизодически появляющихся на виду животных.
На эту биологическую обусловленность реакции в отсутствии стимула, который до этого ее обусловливал, указал еще полвека тому назад советский зоолог и зоопсихолог Д.Н.Кашкаровна примере кошки, которая «стережет мышь и видит, что мышь появляется из норы. Мышь исчезает прежде, чем кошка начнет реагировать. Однако кошка встает и идет к норе. Здесь не может быть вопроса о том, что в момент реакции определяющий стимул отсутствует... Правильный выбор между тремя норами кошка могла бы сделать лишь на основании непосредственно предыдущего появления мыши, т.е. на основе представления о последней»3. Что же касается обезьян, то, как видно из упомянутых исследований, мы явно имеем здесь дело с более сложными комплексными процессами.
Способность высших позвоночных к предметному восприятию, способности к анализу и обобщению, а тем самым к формированию представлений являются важной предпосылкой образования сложных навыков, которые уже рассматривались раньше (см. ч. I) и которые составляют основное содержание накопления индивидуального опыта не только в сенсорной, но и в эффекторной сфере, точнее сказать, в моторно-сенсорной сфере, действующей как единая система.
Общение.
У высших позвоночных особой сложности достигают и процессы общения. Как и у других животных, средства коммуникации включают у них элементы различной модальности — ольфакторные, тактильные.
Ольфакторная сигнализация, т.е. передача информации другим особям химическим путем, преобладает в территориальном поведении, особенно при маркировке местности у хищных и копытных. Для этого служат специальные железы, расположенные в различных участках кожи и выделяющие специфический пахучий секрет. Последний распространяется по воздуху или наносится на различные предметы (стволы и ветки растений, камни и т.д.). Пахучие метки производятся также с помощью испражнений. Выделениями пахучих желез маркируется и след, что способствует нахождению друг друга особями одного вида. Однако во всех этих случаях общение существенно не отличается от того, что мы наблюдаем, скажем, у насекомых.
Видотипичные, инстинктивные компоненты поведения позвоночных, служащие для акустического и оптического общения между животными, как правило, ритуализованы. Оптическое общение осуществляется прежде всего с помо-
3 Кашкаров Д.Н. Современные успехи зоопсихологии. М.; Л., 1928. С. 123—124,
щью выразительных поз и телодвижений, о чем уже говорилось выше. Видоспецифические отличия выражаются часто в малозаметных человеку, но четко генетически фиксированных деталях, вызывающих совершенно специфические, зачастую ритуализованные реакции у особей того же вида. Особенно часто такие различия наблюдаются у близкородственных животных, причем прежде всего в сигнальных системах, связанных с размножением. Здесь они служат одним из факторов биологического разграничения видов.
Конкретные формы оптического общения отличаются у высших позвоночных большим разнообразием и диффе-ренцированностью. Нередко они выражаются в специфических «диалогах» между двумя или несколькими особями, т.е. во взаимной демонстрации частей тела или поз. Особенно это относится к «ритуалам» запугивания или «импонирования». «Импонирование» выражает превосходство одной особи по отношению к другой, имеет и оттенок угрозы и вызова, но, в сущности, не носит агрессивного характера (в отличие от подлинного «запугивания», за которым часто следует нападение). Часто самцы «импонируют» самкам во время «ухаживания» за ними. Подчиненная особь отвечает на «импонирование» позой «покорности», чем окончательно исключается возможность столкновений между животными.
У млекопитающих оптическое общение часто сочетается с ольфакторным (рис. 43), так что выделение систем общения по отдельным модальностям является у этих животных в большой степени условным.
Это относится и к акустическому общению, в качестве примера которого следует, конечно, прежде всего напомнить о криках и песнях птиц. Хотя человеческое ухо неспособно улавливать все издаваемые птицами звуки (значительная часть их находится в ультразвуковом спектре ниже 50 кГц), мы поражаемся огромному разнообразию их голосовых реакций. К последним необходимо добавить и неголосовые звуки, как, например, свист крыльев при полете, постукивание дятла по стволу деревьев и т.д. Видоспецифичность таких звуков делает их пригодными для общения.
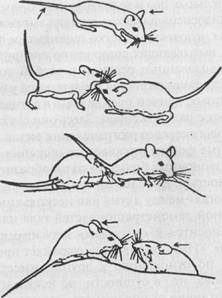
Рис. 43. Оптико-ольфакторное общение у грызунов (род Peromyscus). Стрелками обозначены места взаимного обнюхивания (по Эйзенбергу)
Для биологически адекватного реагирования на все эти звуковые сигналы существенной является видоспецифи-ческая настройка слуховой системы на определенную часть спектра. Как показал советский исследователь биоакустики птиц В.Д.Ильичев, важнейшую роль играют в этой настройке периферические отделы слухового анализатора, выполняющие роль видотипических биологических фильтров. С их помощью осуществляется и специальная «подстройка» слуховой системы на особенно важные для особи звуки. Таким образом, здесь четко выступает характерная для всего инстинктивного поведения взаимообусловленность и единство врожденных функциональных (физиологических и этологических) и морфологических элементов. Зоосемантика звуковых сигналов птиц охватывает практически все сферы их жизнедеятельности. Эти сигналы служат опознаванию особи (в частности, ее видовой принадлежности), оповещению других особей о физиологическом состоянии экспедиента (сюда относится, например, «попрошайничество») или об изменениях в окружающей среде (появление врага или, наоборот, кормового объекта и т.п.). При этом богатые зоопрагматичес-кие средства птицы позволяют экспедиенту передавать перцепиенту весьма детализованную информацию. Так, грач, подлетающий с кормом к гнезду, оповещает об этом насиживающую самку криком, могущим иметь четыре разных оттенка, на которые самка также отвечает разными звуками. Эти оттенки голосовых реакций птиц подчас настолько тонки, что не воспринимаются человеческим ухом.
Дата добавления: 2016-08-07; просмотров: 605;