Часть III ЭВОЛЮЦИЯ ПСИХИКИ. 3 страница
Согласно этой гипотезе, все органы чувств многоклеточных животных развились из органов осязания — наименее дифференцированных рецепторов. В наиболее элементарных случаях осязательная функция присуща всем клеткам поверхности тела. Но уже у кишечнополостных появляются специальные осязательные клетки, которые, скапливаясь в определенных местах, образуют подлинные органы осязания. Это вытянутые цилиндрические или ве-ретеновидные клетки, несущие на конце неподвижный чувствительный волосок или пучок волосков. Однако эти органы часто выполняют и обонятельную функцию. Особенностью низших многоклеточных беспозвоночных является то, что во многих (хотя и не во всех) случаях эти две рецепторные функций слиты и не поддаются морфологическому разграничению.
В этом нетрудно усмотреть остатки первичной плюро-модальности.
С другой стороны, органы зрения относятся к наиболее сложным по строению и функционированию. Иногда органы чувств низшего порядка превращаются в органы чувств высшего порядка (например, у пиявок некоторые из органов осязания — так называемые «сенсиллы» — превращаются в глаза). Известный советский зоолог, специалист по сравнительной анатомии беспозвоночных В.А.Догель говорил в таких случаях о «повышении органа в ранге». Однако в процессе филогенеза, как отмечает Догель, нередко имело место и обратное явление; в других случаях какие-то рецепторы беспозвоночных исчезали, чтобы потом вновь появиться в несколько измененной форме. Это непостоянство и легкость перестройки привели к тому, что у близкородственных беспозвоночных однотипные рецепторы, в частности органы оптической чувствительности, подчас бывают совершенно различными по строению и функциям.
Если взять, к примеру, кишечнополостных, то гидра четко реагирует на свет, хотя специальных органов зрения у нее нет. Она воспринимает свет всей поверхностью тела. Положительный фототаксис гидры выражается в том, что животное производит в освещенной сфере круговые или маятникообразные колебательные движения и в конце концов занимает положение в сторону источника света или даже направляется (ползет) к нему. Свободноживущие же представители кишечнополостных — медузы — обладают уже специальными многоклеточными органами светочувствительности. В простейшем случае эти органы представлены так называемыми глазными пятнами, которые находятся среди обыкновенных эпителиальных клеток, и даже нечетко отграничены от них. Более дифференцированным рецептором является глазная ямка (рис. 32). Однако чаще у медуз встречаются уже настоящие глаза, причем наиболее
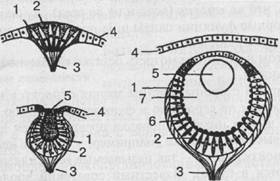
Рис. 32. Развитие глаза беспозвоночных (на примере гидромедуз). Слева вверху — глазное пятно; слева внизу — глазная ямка; справа — глазной пузырь. 1 — зрительная клетка; 2 — пигментная клетка; 3 — зрительный нерв; 4 — эпителий; 5 — хрусталик; 6 — светочувствительные палочки; 7 — стекловидное тело (по Бючли)
сложно устроенные из них представляют собой погруженные под слоем эпителиальных клеток глазные пузыри приблизительно шарообразной формы. Эпителий над глазным пузырем утончен и представляет собой прозрачную роговицу. Дно и стенки пузыря состоят из двух типов клеток: ретинальных и пигментных, причем ретинальные клетки снабжены чувствительными палочками. В полости глазного пузыря находится стекловидное тело - - студенистая масса, защищающая ретину от механических повреждений. Иногда встречаются даже хрусталик и радужка, и тогда налицо все основные компоненты глаза высших животных (нет, однако, глазодвигательных мышц и систем фокусировки).
Учитывая, что глазное пятно является исходной формой вообще всех органов зрения, можно, следовательно, в ряду медуз проследить путь усложнения структуры от самого примитивного органа светочувствительности до сложного, высокодифференцированного глаза.
Очень разнообразны по своему строению и глаза червей, как и других низших многоклеточных беспозвоночных; в принципе к ним относится то же, что говорилось в отношении глаз медуз. В соответствии с многообразием движений кольчатых червей, разнообразием способов добывания пищи и других моментов жизнедеятельности находится и уровень развития сенсорной сферы этих животных. Это, правда, не означает, что у кольчецов имеются рецепторы для всех видов энергии, воздействующих на них, или даже что для всех форм чувствительности имеются специальные органы чувств. Так, например, у кольчатых червей встречаются сложно устроенные глаза, снабженные даже хрусталиками. Имеются весьма сложные глаза и у некоторых планарий и улиток. По их обладатели, насколько известно, неспособны к зрительному восприятию предметов. За исключением, может быть, некоторых улиток, у всех этих животных фотоскопические глаза, позволяющие отличать свет от тьмы и направление, откуда световые лучи падают на животное, а также перемещение светотеней в непосредственной близости от животного. Светочувствительность может при этом быть очень высокой, например, гребешок, двустворчатый моллюск с несколькими десятками глаз, закрывает створки раковины уже при уменьшении интенсивности освещения на 0,3%. (Для сравнения можно указать, что человек воспринимает уменьшение освещения лишь не менее чем на 1%).
Большой интерес представляют активно плавающие многощетинковые кольчатые черви из семейства Alciopidae, ведущие хищный образ жизни. У этих полихет глаз не только отличается исключительно сложным строением и величиной, но и снабжен аккомодационным устройством в виде специальных сократительных волокон, способных передвигать хрусталик и тем самым менять фокусное расстояние. Это единственный известный случай среди низших беспозвоночных: аккомодация глаза встречается только у головоногих моллюсков и позвоночных. Возможно, у этих червей в какой-то степени уже существует предметное зрение, что было бы исключением, подтверждающим общее правило. Это относится и к свободно плавающим хищным моллюскам Heteropoda, которые тоже обладают весьма сложно устроенными глазами с приспособлением, заменяющим аккомодацию.
Что же касается дождевого червя, то здесь обнаруживается чрезвычайно интересный факт: у него нет не только сложно устроенных, но и вообще никаких специальных органов светочувствительности. Вместе с тем ему свойствен четкий отрицательный фототаксис. Функцию све-тоощущения выполняют рассеянные в коже светочувствительные клетки. Это пример кожной светочувствительности низших многоклеточных беспозвоночных. Кожная светочувствительность наблюдается и у многих моллюсков, причем у двустворчатых это нередко единственная форма фоторецепции. Эти моллюски реагируют как на освещение, так и на затемнение чаще всего одинаковым образом -втягиванием выступающих из раковин частей тела или запиранием раковины. Многие улитки реагируют на внезапное затемнение сокращением «ноги», причем эта реакция сохраняется и после экстирпации глаз, что опять-таки указывает на наличие кожной светочувствительности.
Реакция дождевого червя на условия освещения состоит в том, что он уползает в зону большего затемнения. Если же внезапно уменьшить интенсивность освещения, то червь реагирует на это движениями бегства; в естественных условиях он вползает в почву. Такая же реакция следует за внезапным освещением. Если же осветить лишь определенный участок тела дождевого червя, то время этой реакции сокращается пропорционально размеру освещаемой площади поверхности тела — в наибольшей мере червь реагирует при освещении всей его. поверхности. Следовательно, реакция определяется градиентом раздражения освещенных и неосвещенных частей тела. Аналогично реагируют на свет и другие кольчатые черви.
Кольчатые черви реагируют также на прикосновения, химические и термические раздражения, силу тяжести, электрические раздражения, течение воды, а наземные формы (дождевые черви) -- на влажность среды. Однако принципиальных отличий от реакции на свет нет: все эти реакции находятся на одном уровне и характеризуются тем, что являются ответами на отдельные раздражители, на отдельные признаки, качества предметов, но не на сами предметы как таковые. Так, например, много- и малоще-тинковые черви проявляют четко выраженные таксисные реакции на тактильные раздражения. Преобладают отрицательные реакции, но в ряде случаев наблюдаются и положительные тигмотаксисы: прикосновения нервных сегментов животного к субстрату влекут за собой прижимание к нему всем телом, что имеет, конечно, большое значение при роющем образе жизни или жизни в трубках. Интересно, что у дождевого червя соответствующие ре-цепторные образования представлены лишь отдельными чувствительными клетками, разбросанными по всему телу, но более густо располагающимися на его переднем конце. У многощетинковых же червей органами осязания часто являются щупальца или щетинки.
Хорошо развита и химическая чувствительность, причем в большинстве случаев наблюдается отрицательный хемотаксис. При большой интенсивности химического воздействия черви всегда реагируют отрицательно. С другой стороны, дождевой червь, например, способен по химическим признакам выбирать разные виды листьев, что свидетельствует о специализации в сенсорной сфере. У ряда кольчецов обнаружены расположенные около ротового отверстия органы химической чувствительности в виде ямок. Особенно это относится к плавающим видам, у которых имеется пара таких ямок, выстланных мерцательным эпителием.
Таксисы.
Как и на более низких уровнях эволюционного развития, пространственная ориентация совершается на высшем уровне элементарной сенсорной психики, преимущественно на основе примитивных таксисов. Но с усложнением жизнедеятельности организмов возрастают и требования к локализации биологически значимых компонентов среды по их биологически незначимым признакам. Возникает необходимость более сложного таксисного поведения, позволяющего животному достаточно четко и дифференцированно ощущать и реагировать. Именно этим и отличаются животные, находящиеся на рассматриваемом здесь уровне развития психики: благодаря симметричному расположению органов чувств у них обнаруживаются наряду с кинезами и элементарными таксисами и некоторые высшие формы таксисного поведения,
Немецкий ученый А.Кюн выделил следующие категории высших таксисов, которые, правда, вполне развиты лишь у высших животных: тропотаксисы — движение ориентируется по равнодействующей, образуемой в результате выравнивания интенсивности возбуждения в симметрично расположенных рецепторах; телотаксисы — выбор и фиксация одного источника раздражения и направление движения к этому источнику («цели»); менотаксисы — при несимметричном раздражении в симметрично расположенных рецепторах движение производится под углом к источнику раздражения. Менотаксисы играют ведущую роль в сохранении животным константного положения в пространстве.
Кроме того, у высших животных с развитой памятью встречается еще одна форма таксисов — мнемотаксисы, при которых основную роль играет индивидуальное запоминание ориентиров, что особенно важно для территориального поведения, о чем еще пойдет речь.
В поведении кольчецов тропо- и телотаксисы чаще всего проявляются совместно: если животное подвергнуть одновременному воздействию двух источников энергии (например, света) одинаковой силы, то при положительном таксисе оно будет первоначально двигаться по направлению равнодействующей, т.е. линии, ведущей к середине расстояния между источниками (тропотаксис), но затем, как правило, еще не дойдя до этой точки, оно повернет и направится к одному из источников энергии (телотаксис).
Наиболее четко тропо- и телотаксисы проявляются у активно двигающихся кольчатых червей, прежде всего у хищников. Последние не только обнаруживают присутствие жертвы по исходящим от нее физическим и химическим стимулам, но и направляются к ней, ориентируясь на основе таких таксисов. Наземные пиявки, например, способны точно локализовать местонахождение жертвы и с удивительной быстротой направляются к ней, ориентируясь по сотрясению субстрата, а затем, на более близком расстоянии, и по исходящему от жертвы (крупных млекопитающих) теплу и запаху. В результате хищник обнаруживает ее по соответствующим градиентам с помощью сочетания положительных вибрационных, термических и химических телотаксисов.
При менотаксисах животное, как говорилось, двигается под углом к линии воздействия, исходящей от источника энергии. Типичный пример — ориентация мигрирующих животных по солнечному (или другому астрономическому) «компасу», что, конечно, особенно характерно для перелетных птиц. У низших многоклеточных беспозвоночных менотаксисы обнаружены у некоторых улиток. Если этих животных поместить на освещаемый сбоку и медленно вращающийся диск, то они, двигаясь против направления вращения, придерживаются определенного угла по отношению к источнику света. В этих случаях, следовательно, можно уже говорить об ориентации животного по световому компасу, которая играет исключительную роль в жизни высших животных.
Зачатки высших форм поведения.
Многощетинковые кольчатые черви относятся к наиболее развитым в психическом отношении низшим беспозвоночным. Их поведение отличается подчас большой сложностью и представляет особый интерес в том смысле, что содержит ряд элементов психической деятельности, присущей более высокоорганизованным животным. В отличие от основной массы низших беспозвоночных у полихет наблюдаются некоторые существенные усложнения видотипичного поведения, отчасти уже выходящие за рамки типичной элементарной сенсорной психики.
Так, в некоторых случаях у этих морских червей встречаются действия, которые уже можно назвать конструктивными, поскольку животные активно создают сооружения из отдельных посторонних частиц, скрепляя их в единое целое. Речь идет о постройке уже упомянутых «домиков»-трубок из отдельных частичек, которые собираются червями на морском дне и укрепляются с помощью специальных «рабочих» органов — преобразованных передних параподий.
Сам процесс сооружения трубки является сложной деятельностью, состоящей из нескольких фаз, адекватно видоизменяющихся в зависимости от таких внешних факторов, как характер грунта и течения, рельеф дна, количество и состав опускающихся на дно частиц, скорость их оседания и т.д. Строительным материалом служат разного рода мелкий обломочный материал (например, кусочки раковин), песчинки, частицы растений и т.п., которые скрепляются специальными выделениями особого «цементирующего органа». Пригодные для употребления частицы выбираются животным, причем в зависимости от его возраста; молодые особи используют лишь мелкие гранулы, более старые — более крупные. Все частицы собираются и прикрепляются друг к другу уже упомянутыми специализированными щупальцами. Полихета Aulophorus carteri, например, строит трубки из спор водных растений, которые она собирает и прикрепляет друг к другу вокруг себя подобно тому, как строится труба из кирпичей (рис. 33).
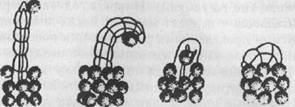
Рис. 33. Постройка многощетинковым червем трубки из спор водных растений (по Грассэ)
Не меньший интерес представляют впервые намечающиеся у многощетинковых червей проявления брачного поведения и агрессивности, а вместе с ними и элементов общения. Конечно, подлинные агрессивность и брачное поведение характеризуются ритуализацией, которая появляется только у головоногих моллюсков и членистоногих, т.е. на низшем уровне перцептивной психики. Все же у представителей рода Nereis удалось наблюдать борьбу между двумя червями. У Nereis pelagica такая борьба может начаться (но не обязательно), если одна особь пытается проникнуть в «домик» другой особи. Иногда вторгшаяся особь кусает владельца «домика» в задний конец тела, и тогда «хозяин» покидает трубку или же, перевернувшись, вступает в борьбу с пришельцем. В других случаях, когда животные встречаются в трубке на полпути, головой к голове, борьба вспыхивает немедленно, и черви борются, расположившись брюшными сторонами и головами друг к другу. Однако при всех обстоятельствах животные не наносят друг другу повреждения. В более ожесточенной, «непримиримой» форме борьба происходит у Nereis caudata, причем не только из-за обладания «домиком». «Драки» описаны английским исследователем поведения морских кольчатых червей С.М.Эвансом и его сотрудниками также у полихеты Harmothoe imbricata. У этого вида борьба может возникнуть при случайной встрече двух особей, особенно же агрессивность проявляется в сфере воспроизведения, при образовании пар: самец, уцепившийся за самку, становится чрезвычайно агрессивным по отношению к другим самцам (но не самкам). Никогда, однако, борьба не сопровождается у полихет подачей каких-либо сигналов или другими проявлениями ритуализации поведения.
Виноградные улитки же выполняют сложные «брачные игры», длящиеся иногда по нескольку часов, во время которых партнеры принимают по отношению друг к другу различные позы, колют друг друга известковыми спикулами («любовными стрелами») и т.д. Только после такой взаимной стимуляции начинается собственно спаривание (перенос сперматофора). У некоторых полихет, например у Platynereis dumerilii, в последние годы также были описаны брачные «танцы», однако, очевидно, еще рано делать выводы о их конкретном значении и о том, имеют ли они какое-то отношение к ритуализованному поведению. Но во всех этих случаях, несомненно, обнаруживаются некие зачатки, предваряющие значительно более сложные формы поведения высших позвоночных.
Пластичность поведения.
Поведение кольчатых червей, как и других низших беспозвоночных, характеризуется малой пластичностью, консервативностью. Господствуют врожденные стереотипы («врожденные программы поведения»). Индивидуальный опыт, научение, играет в жизни этих животных еще небольшую роль. Ассоциативные связи формируются у них с трудом и лишь в ограниченных пределах. Результаты научения сохраняются недолго. У всех кольчатых червей встречается наиболее простая форма научения — привыкание, с которым мы уже встречались у простейших, у которых она является главной, если не единственной формой модификации врожденного, видотипичного поведения. У низших многоклеточных животных врожденные реакции на определенные раздражения также прекращаются после многократного повторения, если не последует адекватного подкрепления этих реакций. Так, например, дождевые черви перестают реагировать на повторное затенение, если оно остается без последствий.
Отмечается привыкание и в сфере пищевого поведения: если многощетинкового червя повторно «кормить» комочками бумаги, смоченными соком его обычной жертвы, он перестает их принимать. Если попеременно с такими комочками давать ему подлинные кусочки пищи, то он в конце концов научается различать их и будет отвергать лишь несъедобную бумагу. Подобные опыты ставились и на кишечнополостных (на полипах, находящихся, как мы уже знаем, на более низком уровне элементарной сенсорной психики). При этом полипы вели себя так же, как морские черви: уже после нескольких (до 5) опытов они отбрасывали несъедобные объекты еще до того, как подносили их к ротовому отверстию. Интересно, что нетренированные щупальца этого не делали, даже находясь рядом с тренированными.
Эти эксперименты интересны тем, что показывают способность низших беспозвоночных отличить по побочным физическим качествам съедобное от несъедобного (по вкусу предлагавшиеся объекты были одинаковыми), что подтверждает наличие истинного психического отражения уже на этом, низшем, уровне филогенетического развития. Ведь здесь производится опосредствованное действие, отличающееся тем, что свойство (или сочетание нескольких свойств), которым животное руководствуется при оценке пригодности объекта к пищевому употреблению, выступает как подлинный сигнал, а чувствительность червя или полипа играет роль посредника между организмом и компонентом среды, от которого непосредственно зависит существование животного.
Более сложное научение путем «проб и ошибок» и формирования новой индивидуальной двигательной реакции можно в элементарной форме обнаружить, начиная уже с плоских червей. Так, планарии, встречая на своем пути полоску наждачной бумаги, сперва останавливаются, но затем все же переползают через нее. Если же сочетать соприкосновение червя с наждачной бумагой с его сотрясением, то планарии перестают переползать через нее и тогда, когда не производится сотрясения. Правда, здесь, очевидно, еще нет подлинной ассоциации между двумя раздражителями (шероховатость и сотрясение); скорее всего, происходит суммация первоначально не очень сильного отрицательного раздражения (шероховатость) с дополнительным отрицательным раздражением (сотрясение) и что, вероятно, влечет за собой общее повышение возбудимости животного.
Все же можно выработать уже у планарии и более сложные реакции, когда один из раздражителей является биологически «нейтральным». В таких случаях можно говорить, очевидно, уже об элементарных процессах подлинного ассоциативного научения. Так, например, Л.Г.Воронин и Н.А.Тушмалова сумели выработать у планарии и кольчатых червей оборонительные и пищевые условные рефлексы. При этом были получены и данные, свидетельствующие об усложнении форм временных связей. Если по поводу плоских червей можно говорить лишь о примитивных неустойчивых условных рефлексах, то у многощетинковых червей обнаруживаются уже стабильные самовосстанавливающиеся после угасания условные рефлексы. Эти различия соответствуют глубоким морфологическим различиям в строении нервной системы этих групп животных и свидетельствуют о существенном прогрессе психической активности у полихет.
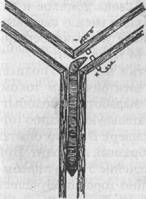
Пластичность поведения дождевых червей очень убедительно была показана еще в 1912 г. известным американским зоопсихологом Р.Иерксом. В проведенных им опытах черви должны были в Т-образном лабиринте выбирать определенную сторону, где находилось «гнездо» (в противоположной стороне червь получал электрический удар). Чтобы этому научить червей, потребовалось 120— 180 опытов. (Улитки осваивают такую задачу после 60 опытов и запоминают верное решение в течение 30 дней.) Впоследствии аналогичные опыты с дождевыми червями ставились Л.Геком, И.С.Робинсоном, Р.И.Войтусяком (рис. 34) и другими исследователями. Удавалось также переучивать червей, поменяв «гнездо» и электроды местами. Характерно, что результат научения не меняется и после удаления передних сегментов. Более того, даже лишенный с самого начала этих сегментов, червь оказывается способным научиться правильно ориентироваться в лабиринте.
|
Рис. 34. Научение у дождевого червя (опыты Войтусяка)
Подобные факты побудили в свое время В.А. Вагнера говорить о «сегментарной психологии» кольчатых червей и других «членистых», имея при этом в виду, что ганглии каждого сегмента в большой степени обеспечивают автономное выполнение элементарных психических функций. Обезглавленные кольчецы, писал Вагнер, «не теряют способности к спонтанным движениям, за исключением тех лишь, которые стоят в прямой зависимости и связи с органами чувств головы... Обезглавленные черви удерживают свои инстинктивные действия, даже те, в которых голова принимает прямое участие»2.
Все это свидетельствует о том, что на уровне даже высших червей цефализация еще не достигла такого развития, чтобы оказать решающее влияние на все поведенческие акты животного. Все же общая направляющая роль головного мозга достаточно велика уже и на этом уровне. Это проявляется, например, в том, что если разрезать пополам дождевого червя, передняя половина будет закапываться в почву, производя вполне координированные движения, задняя же будет лишь беспорядочно извиваться.
Еще лучше, чем у малощетинковых червей, как уже отмечалось, вырабатываются ассоциативные связи у по-
2 Вагнер В.А. Сегментарная психология. Новые идеи в биологии. Сборник № 6. Биопсихология. Спб., 1914. С. 123.
лихет. В опытах удавалось, например, переделать отрицательную реакцию на освещение в положительную путем сочетания освещения с подкармливанием. В итоге черви стали при освещении выползать из своих домиков и без пищевого подкрепления. Время реакции (между световым раздражением и ответным движением) уменьшилось при этом с 28,9 до 3,5 секунды (опыты М.Купленда).
Обстоятельно изучал способность к научению полихет С.М.Эванс. Применяя наказания электрическим током, этот исследователь довольно легко выработал у подопытных животных отрицательную двигательную реакцию (отказ от вползания в узкий темный коридор). В других опытах червям приходилось проползать Т-образный лабиринт. При правильном повороте они могли в течение 5 минут пребывать в затемненной камере. Особенно хорошо обучались полихеты Nereis virens, которым понадобилось менее 100 сочетаний для почти безошибочного решения задачи и ее прочного запоминания; спустя 48 часов они безошибочно воспроизводили правильный путь через лабиринт (рис. 35).
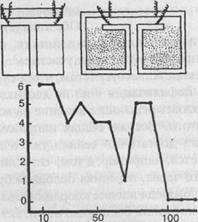
Рис. 35. Научение у Nereis virens (опыт Эванса). Вверху даны схемы Т-образных лабиринтов, в которых производились опыты. Внизу — кривая научения (по оси ординат — число ошибок, по оси абсцисс — число опытов)
Общая характеристика высшего уровня элементарной сенсорной психики.
Как уже отмечалось, наиболее низкоорганизованные формы многоклеточных беспозвоночных стоят на том же уровне психического развития, что и высшие представители простейших. Но то, что здесь не обнаруживаются существенные различия в поведении, несмотря на глубокие различия в строении, не должно нас удивлять, ибо, как уже говорилось, простейшие олицетворяют собой совершенно особую, рано отклонившуюся филогенетическую ветвь, которая до известных пределов развивалась параллельно ветви низших многоклеточных животных.
Что же касается поведения кольчатых червей, то оно вполне отвечает стадии элементарной сенсорной психики, ибо слагается из движений, ориентированных лишь по отдельным свойствам предметов и явлений (или их сочетаниям), причем это те свойства, которые, по Леонтьеву, оповещают о появлении жизненно важных условий среды, от которых зависит осуществление основных биологических функций животных. Эта ориентация осуществляется, таким образом, на основе одних лишь ощущений. Перцепция, способность к предметному восприятию, еще отсутствует. Не исключено, правда, что у некоторых наземных улиток, как и у упомянутых выше свободно плавающих хищных моллюсков и полихет, уже намечаются зачатки этой способности. Так, виноградная улитка обходит преграду еще до прикосновения к ней, ползет вдоль нее, но только если преграда не слишком велика; если же изображение преграды занимает всю сетчатку, улитка наталкивается на нее. Не реагирует она и на слишком мелкие предметы.
В поведении кольчатых червей еще преобладает избегание неблагоприятных внешних условий, как это имеет место у простейших. Но все же активный поиск положительных раздражителей занимает в поведении кольчецов уже заметное место, и это весьма характерно для высшего уровня элементарной сенсорной психики. Так же как и у простейших, в жизни кольчатых червей и других низших многоклеточных беспозвоночных большую роль играют кинезы и элементарные таксисы. Но наряду с ними здесь уже встречаются зачатки сложных форм инстинктивного поведения (особенно у некоторых многощетинковых червей, пиявок, а также улиток) и впервые появляются высшие таксисы, обеспечивающие значительно более точную и экономную ориентацию животного в пространстве, а тем самым и более полноценное использование пищевых ресурсов в окружающей среде. В результате возникли предпосылки для поднятия всей жизнедеятельности на более высокую ступень, что и имело место на стадии перцептивной психики.
Как было показано, у высших представителей рассматриваемой группы беспозвоночных впервые появляются зачатки конструктивной деятельности, агрессивного поведения, общения. Здесь опять подтверждается правило, что высшие формы поведения зарождаются уже на низших стадиях развития психической деятельности.
Давая общую оценку поведению низших многоклеточных беспозвоночных, следует отметить, что, очевидно, первично главная функция еще примитивной нервной системы состояла в координации внутренних процессов жизнедеятельности в связи со все большей специализацией клеток и новых образований — тканей, из которых строятся все органы и системы многоклеточного организма. «Внешние» же функции нервной системы определяются степенью внешней активности, которая у этих животных находится на еще невысоком уровне, зачастую не более высоком, чем у высших представителей простейших. Вместе с тем строение и функции рецепторов, как и «внешняя» деятельность нервной системы, значительно усложняются у животных, ведущих более активный образ жизни. Особенно это относится к свободноживущим, активно передвигающимся формам.
Говоря об обширной и пестрой по составу группе низших беспозвоночных, нужно, конечно, учесть, что поведение этих животных еще очень слабо изучено. Мы, например, еще почти ничего не знаем об онтогенезе поведения этих животных, о том, как формируется и развивается их поведение, да и совершенствуется ли оно вообще в процессе индивидуального развития, на что уже указывалось в части, посвященной онтогенезу поведения. Вполне возможно (если, конечно, исключить метаморфозные преобразования, личиночное поведение у низших многоклеточных и т.п.), что подобное онтогенетическое совершенствование у обсуждаемых животных не является существенным или даже вообще не происходит, ибо жесткость врожденных программ поведения, исключительная стереотипия форм реагирования являются у этих животных определяющей чертой всего их поведения.
Дата добавления: 2016-08-07; просмотров: 620;