Часть III ЭВОЛЮЦИЯ ПСИХИКИ. 2 страница
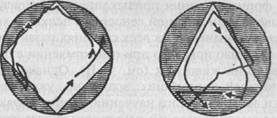
Рис. 30. Привыкание у парамеции (по Брамштедту). Пояснения см. в тексте
Все же, очевидно, мы имеем здесь дело с элементарной формой научения - привыканием. Привыкание (к измененным внешним условиям), как мы уже знаем, играет немаловажную роль и в поведении высших животных, но имеет качественно иной характер уже хотя бы потому, что строится у них не только на ощущениях, но и на восприятиях. При элементарной же сенсорной психике животное может привыкать лишь к воздействию отдельных раздражителей (или их совокупностей), являющихся воплощением отдельных свойств или качеств компонентов окружающей среды. Именно это и имело место в приведенных примерах, когда наблюдалась модификация ви-дотипичного поведения: врожденные реакции на определенные раздражители постепенно элиминируются, если при многократном повторении не наступит биологически значимый эффект. Так, в опытах Тушмаловой фобическая реакция у инфузорий со временем сводилась на нет при повторной встрече с «безобидным» раздражителем.
Привыкание необходимо отличать от утомления, выражающегося в явлениях «истощения» животного. В опытах Тушмаловой это выражалось в неспособности инфузории к дальнейшему реагированию, если она в течение 2—3 часов подвергалась очень сильным раздражениям. Утомление связано с перерасходом энергетических ресурсов, привыкание же представляет собой активную приспособительную реакцию, значение которой состоит в экономии этих ресурсов, в предотвращении траты энергии на бесполезные для животного движения.
Как форма научения привыкание характеризует низший уровень элементарной сенсорной психики, хотя и не теряет своего значения на всех ступенях развития психики, как это было показано при ознакомлении с онтогенезом поведения животных (см. ч. II). Однако у высших представителей простейших, возможно, уже существуют и зачатки ассоциативного научения, вообще характерного для более высоких уровней психического развития.
Об ассоциативном научении можно говорить в тех случаях, когда устанавливается временная связь между биологически значимым и «нейтральным», точнее, биологически маловалентным раздражителем. Именно такую связь Брам-штедту удалось выработать у туфельки, которая, как уже отмечалось, не реагирует заметным образом на изменение освещения, но весьма чувствительна к температурным изменениям. Если в эксперименте затенить одну половину капли воды, в которой плывут парамеции, и при этом подогреть освещенную часть капли, то вскоре инфузории соберутся в холодной темной части, но останутся там (в течение 15 минут) и после того, как температура в обеих частях капли будет уравновешена. Правда, эти опыты Брамштедта подвергались серьезной критике, так как при подогреве меняется и химизм воды (меняется растворимость содержащихся в ней газов), что не может не влиять на поведение инфузории (опыты У.Грабовского).
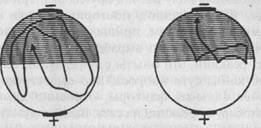
Рис. 31. Образование у парамеции временной связи между электрическим раздражением и светом (по Засту). Пояснения см. в тексте
Вместе с тем аналогичный результат может быть получен и путем наказания животных при заплыве в освещенную зону ударами электрического тока (рис. 31). Эффект избегания сохранялся в этом случае в течение 20 минут (опыты Г.Зеста). Однако обратный результат, т.е. избегание темноты и заплыв в освещенную зону, Зесту получить не удалось. Это обстоятельство, однако, является результатом методической ошибки: инфузориям свойствен гальванотаксис, выражающийся в их устремлении к катоду. В упомянутых же опытах Зеста катод всегда находился на темной половине и гальванотаксис оказывался сильнее электрического раздражения, почему инфузории и направлялись только в темноту.
В более тщательно поставленных опытах польскому ученому С.Вавржинчику удалось преодолеть эти методические недостатки и с успехом научить инфузории избегать темный участок стеклянной трубки, где их раздражали электрическим током. Постепенно туфельки все чаще оставались в освещенной зоне и поворачивали на границе с затемненной зоной даже при перемещении этой границы вдоль трубки еще до получения электрического удара. Наконец, они оставались в освещенной зоне в четыре раза дольше, чем в темной, даже тогда, когда раздражения током совсем прекратились. Эта реакция сохранялась даже в течение 50 минут, что является достаточно большим сроком для простейших. Четкие положительные результаты этот исследователь получил и в других экспериментах.
Правда, Я.Дембовский, повторивший эти опыты с некоторыми изменениями, пришел к выводу, что, хотя и не исключена возможность выработки у простейших обусловленных реакций, эти опыты с туфельками все-таки не решают до конца сути вопроса, ибо существуют несравненно более сильные факторы, особенно химические, которые маскируют реакции на свет. Ввиду отсутствия вполне убедительных экспериментальных данных Дембовский оставил вопрос о наличии у простейших обусловленных реакций открытым.
Вместе с тем и у других (кроме туфельки) инфузорий, в частности у стентора и сувойки, удалось после 140—160 сочетаний выработать сократительные реакции в ответ на световое раздражение.
На инфузориях проводились и другие опыты с применением разных методических приемов. В частности, использовались капилляры, иногда с загнутым концом, в которые сажались парамеции, после, чего измерялось время, необходимое им для высвобождения. В новейших исследованиях, проведенных таким способом, получались вполне положительные результаты: при каждом повторении опыта время выхода парамеции из трубочки заметно уменьшалось, что и признается доказательством их способности к ассоциативному научению.
Однако американские ученые Ф.Б.Эпплуайт и Ф.Т.Гарднер, недавно повторившие эти опыты, считают такой вывод необоснованным. Эти ученые засасывали последовательно в один и тот же капилляр разные, еще ни разу не побывавшие в опыте туфельки и установили, что время выхода этих животных из капилляра также постепенно уменьшается. Но этого не случалось, если капилляр до употребления тщательно промывался и кипятился в деио-низированной воде. Использование обработанных таким образом капилляров для повторного проведения опыта с одной и той же туфелькой не давало практически никакого эффекта научения. Отсюда экспериментаторы вывели заключение, что уменьшение времени выхода из капилляра является у парамеции не результатом научения, а реакцией на загрязнение внутренней поверхности трубки продуктами обмена веществ, которое увеличивается от опыта к опыту и от которого животные пытаются все скорее уйти.
С другой стороны, в аналогичных исследованиях других ученых учитывались подобные возможные изменения среды, и было установлено, что время выхода инфузорий из трубки сокращается лишь в первых опытах, потом же оно остается постоянным (среда в трубке должна, однако, продолжать изменяться).
Таким образом, вопрос о наличии у инфузорий (а тем более у простейших вообще) ассоциативного научения еще нельзя считать решенным. Все же, очевидно, такое научение существует у них в зачаточном виде.
Общая характеристика психической активности.
Как мы могли убедиться, на низшем уровне элементарной сенсорной психики поведение животных выступает в достаточно разнообразных формах, но все же мы имеем здесь дело лишь с примитивными проявлениями психической активности. О такой активности, о психике мы можем здесь говорить потому, что простейшие активно реагируют на изменения в окружающей их среде, причем реагируют на биологически непосредственно не значимые свойства компонентов среды как на сигналы о появлении жизненно важных условий среды. Другими словами, простейшим свойственна элементарная форма психического отражения -- ощущение, т.е. чувствительность в собственном смысле слова. Но там, как мы уже знаем, где появляется способность к ощущению, начинается психика.
Уже говорилось и о том, что даже низший уровень психического отражения не является низшим уровнем отражения вообще, существующего в живой природе: растениям присуще допсихическое отражение, при котором имеют место лишь процессы раздражимости. Однако элементы такого допсихического отражения встречаются и у простейших.
Большой интерес представляют в этом отношении, как и вообще для понимания условий зарождения психического отражения, реакции простейших на температуру. Здесь непосредственно необходимая для поддержания жизни энергия еще тождественна опосредующей энергии, сигнализирующей о наличии жизненно необходимого компонента среды.
Еще в начале нашего века М.Мендельсон установил, что у инфузорий реакции на изменения температуры становятся все более дифференцированными по мере приближения к некому термическому оптимуму. Так, у Paramecium aurelia такой оптимум находится в пределах +24 ... +28° температуры воды. При +6 ... +15° инфузория реагирует на разности температуры от 0,06 до 0,08°, при +20 ... +24° же — на разности от 0,02 до 0,005°. Но по современным данным этот вид хорошо переносит и низкие температуры и даже замораживание до -30° и более.
Вместе с тем у простейших специфические терморецепторы, т.е. специальные образования, служащие для восприятия изменений температуры среды, не обнаружены. (Достоверно такие рецепторы неизвестны и для других беспозвоночных.) В свое время Г.Дженнингс полагал, что у инфузории (туфельки) передний конец тела особенно чувствителен в отношении температурных раздражений. Однако впоследствии О.Кёлер показал, что у разрезанной на две части туфельки передний и задний концы тела ведут себя в этом отношении в принципе так же, как целое животное. Отсюда напрашивается вывод, что реакция на изменения температуры является у простейших свойством всей протоплазмы. Возможно, что реакции простейших на термические раздражения еще сходны с биохимическими реакциями типа ферментативных процессов.
Очевидно, все это и соответствует тому обстоятельству, что для организма простейшего тепловая энергия еще не расчленяется на источник жизнедеятельности и сигнал о наличии такого источника. В связи с этим отсутствуют и специальные терморецепторы. Мы имеем, следовательно, здесь пример «сосуществования» у простейших допсихического и психического отражения.
Интересно, что отсутствие терморецепторов не делает реакции простейших менее выразительными, чем их реакции на другие внешние агенты. Как было показано, эти реакции проявляются в таксисах (в данном случае термотаксисах) и кинезах.
Еще явственнее «сосуществование» допсихического и психического отражения выступает у низших представителей типа. У эвглены оно обусловлено и наличием наряду с животным, растительного, аутотрофного типа питания, почему эвглена в одинаковой степени относится и к растениям и к животным.
С другой стороны, мы знаем, что качества психического отражения определяются тем, насколько развиты способности к движению, пространственно-временной ориентации и к изменению врожденного поведения. У простейших мы встречаем разнообразные формы передвижения в водной среде, но только на самом примитивном уровне инстинктивного поведения — кинезов. Ориентация поведения осуществляется только на основе ощущений и ограничена элементарными формами таксисов, позволяющими животному избегать неблагоприятные внешние условия. За отдельными исключениями, активность простейших находится в целом как бы под отрицательным знаком, ибо эти животные попадают в сферу действия положительных раздражителей, уходя от отрицательных.
Это значит, что поисковая фаза инстинктивного поведения (кинезов) в указанном отношении еще крайне недоразвита. К тому же она явно лишена сложной, многоэтапной структуры. Возможно, что во многих случаях эта фаза вообще отсутствует. Во всем этом проявляется не только исключительная примитивность инстинктивного поведения на данном уровне, но и предельная скудность содержания психического отражения. Ведь это содержание наполняется прежде всего активным поиском и оценкой раздражителей на первых этапах поисковой фазы.
Как уже отмечалось, в отдельных случаях у простейших встречаются и положительные элементы пространственной ориентации. К уже упомянутым примерам положительных таксисов можно добавить, что амеба в состоянии находить пищевой объект на расстоянии до 20— 30 микрон. Зачатки активного поиска жертвы существуют, очевидно, и у хищных инфузорий. Однако во всех этих случаях положительные таксисные реакции еще не носят характера подлинного поискового поведения, поэтому эти исключения не меняют общую оценку поведения простейших, а тем более характеристику низшего уровня элементарной сенсорной психики в целом: дистантно на этом уровне распознаются преимущественно отрицательные компоненты среды; биологически «нейтральные» же признаки положительных компонентов, как правило, еще не воспринимаются на расстоянии как сигнальные, т.е. попросту еще не существуют для животного как таковые. Таким образом, психическое отражение выполняет на самом низком уровне своего развития в основном сторожевую функцию и отличается поэтому характерной «однобокостью»: сопутствующие биологически не значимые свойства компонентов среды дистантно ощущаются животными как сигналы появления таких компонентов лишь в том случае, если эти компоненты являются для животных вредными. Что касается, наконец, пластичности поведения простейших, то и здесь простейшие обладают лишь самыми элементарными возможностями. Это вполне закономерно: элементарному инстинктивному поведению может соответствовать лишь элементарное научение. Последнее, как мы видели, представлено наиболее примитивной формой — привыканием, и лишь в отдельных случаях, может быть, встречаются зачатки ассоциативного научения.
Конечно, при всей своей примитивности поведение простейших является все же достаточно сложным и гибким, во всяком случае в тех пределах, которые необходимы для жизни в своеобразных условиях микромира. Эти условия отличаются рядом специфических особенностей, и этот мир нельзя себе представить как просто во много раз уменьшенный макромир. В частности, среда микромира является менее стабильной, чем среда макромира, что проявляется, например, в периодическом высыхании маленьких водоемов. С другой стороны, непродолжительность жизни микроорганизмов как отдельных особей (частая смена поколений) и относительное однообразие этого микромира делают излишним развитие более сложных форм накопления индивидуального опыта. В этой микросреде нет столь сложных и разнообразных условий, к которым можно приспособиться только путем научения. В таких условиях пластичность самого строения простейших, легкость образования новых морфологических структур в достаточной мере обеспечивают приспособляемость этих животных к условиям существования. Можно, очевидно, сказать, что пластичность поведения здесь еще не превзошла пластичность строения организма.
Как уже отмечалось, простейшие не являются однородной группой животных, и различия между их разными формами очень велики. Высшие представители этого типа во многих отношениях развивались в своеобразных формах неклеточного строения параллельно низшим многоклеточным беспозвоночным животным. В результате высокоразвитые простейшие проявляют подчас даже более сложное поведение, чем некоторые многоклеточные беспозвоночные, также стоящие на низшем уровне элементарной сенсорной психики. Это тоже одна из причин, почему мы описали этот уровень психического развития на примере только простейших. Здесь наглядно выступает уже отмеченная общая закономерность: психологическая классификация не вполне совпадает с зоологической, так как некоторые представители одной и той же таксономической категории могут еще находиться на более низком психическом уровне, другие - - уже на более высоком. Именно последнее имеет место у высших представителей типа простейших, которых в этом отношении можно было бы рассматривать как исключения. Однако, в сущности, это не так, ибо здесь проявляется и другая закономерность эволюции психики, а именно: элементы более высокого уровня психического развития всегда зарождаются в недрах предшествующего, более низкого уровня. В данном случае, например, примитивные формы ассоциативного научения, вообще характерные для более высокого уровня элементарной сенсорной психики, встречаются в зачатке уже у некоторых видов, относящихся к типу, который в целом стоит на низшем уровне элементарной сенсорной психики, где типичной формой индивидуально-изменчивого поведения является привыкание.
Высший уровень развития элементарной сенсорной психики.
Высшего уровня элементарной сенсорной психики достигло большое число многоклеточных беспозвоночных. Однако, как отмечалось, часть низших многоклеточных беспозвоночных находится в основном на том же уровне психического развития, что и многие простейшие. Это относится прежде всего к большинству кишечнополостных и к низшим червям, а тем более к губкам, которые во многом еще напоминают колониальные формы одноклеточных (жгутиковых). Неподвижный, сидячий образ жизни взрослых губок привел даже к редукции их внешней активности, поведения (при полном отсутствии нервной системы и органов чувств). Но даже у самых примитивных представителей многоклеточных животных создались принципиально новые условия поведения в результате появления качественно новых структурных категорий — тканей, органов, систем органов. Это и обусловило возникновение специальной системы координации деятельности этих многоклеточных образований и усложнившегося взаимодействия организма со средой — нервной системы.
К низшим многоклеточным беспозвоночным относятся помимо уже упомянутых еще иглокожие, высшие (кольчатые) черви, отчасти моллюски и др.
Мы рассмотрим в дальнейшем в качестве примера кольчатых червей, у которых в полной мере выражены признаки поведения, характерные для высшего уровня элементарной сенсорной психики. К кольчатым червям относятся живущие в морях многощетинковые черви (полихеты), малощетинковые черви (наиболее известный представитель — дождевой червь) и пиявки. Характерным признаком строения кольчецов является внешняя и внутренняя метамерия: тело состоит из нескольких большей частью идентичных сегментов, каждый из которых содержит «комплект» внутренних органов, в частности пару симметрично расположенных ганглиев с нервными коммисурами. В результате нервная система кольчатых червей имеет вид «нервной лестницы».
Нервная система.
Как известно, нервная система впервые появляется у низших многоклеточных беспозвоночных. Возникновение нервной системы — важнейшая веха в эволюции животного мира, и в этом отношении даже примитивные многоклеточные беспозвоночные качественно отличаются от простейших. Важным моментом здесь является уже резкое ускорение проводимости возбуждения в нервной ткани: в протоплазме скорость проведения возбуждения не превышает 1—2 микрон в секунду, но даже в наиболее примитивной нервной системе, состоящей из нервных клеток, она составляет 0,5 метра в секунду!
Нервная система существует у низших многоклеточных в весьма разнообразных формах: сетчатой (например, у гидры), кольцевой (медузы), радиальной (морские звезды) и билатеральной. Билатеральная форма представлена у низших (бескишечных) плоских червей и примитивных моллюсков (хитон) еще только сетью, располагающейся вблизи поверхности тела, но выделяются более мощным развитием несколько продольных тяжей. По мере своего прогрессивного развития нервная система погружается под мышечную ткань, продольные тяжи становятся более выраженными, особенно на брюшной стороне тела. Одновременно все большее значение приобретает передний конец тела, появляется голова (процесс цефализации), а вместе с ней и головной мозг — скопление и уплотнение нервных элементов в переднем конце. Наконец, у высших червей центральная нервная система уже вполне приобретает типичное строение «нервной лестницы», при котором головной мозг располагается над пищеварительным трактом и соединен двумя симметричными коммису-рами («окологлоточное кольцо») с расположенными на брюшной стороне подглоточными ганглиями и далее с парными брюшными нервными стволами. Существенными элементами являются здесь ганглии, поэтому говорят и о ганглионарной нервной системе, или о «ганглионарной лестнице». У некоторых представителей данной группы животных (например, пиявок) нервные стволы сближаются настолько, что получается «нервная цепочка».
От ганглиев отходят мощные проводящие волокна, которые и составляют нервные стволы. В гигантских волокнах нервные импульсы проводятся значительно быстрее благодаря их большому диаметру и малому числу синапти-ческих связей (мест соприкосновения аксонов одних нервных клеток с дендритами и клеточными телами других клеток). Что же касается головных ганглиев, т.е. мозга, то они больше развиты у более подвижных животных, обладающих и наиболее развитыми рецепторными системами. Зарождение и эволюция нервной системы обусловлены . необходимостью координации разнокачественных функциональных единиц многоклеточного организма, согласования процессов, происходящих в разных частях его при взаимодействии с внешней средой, обеспечения деятельности сложно устроенного организма как единой целостной системы. Только координирующий и организующий центр, каким является центральная нервная система, может обеспечить гибкость и изменчивость реакции организма в условиях многоклеточной организации.
Огромное значение имел в этом отношении и процесс цефализапии, т.е. обособления головного конца организма и сопряженного с ним появления головного мозга. Только при наличии головного мозга возможно подлинно централизованное «кодирование» поступающих с периферии сигналов и формирование целостных «программ» врожденного поведения, не говоря уже о высокой степени координации всей внешней активности животного.
Разумеется, уровень психического развития зависит не только от строения нервной системы. Так, например, близкие к кольчатым червям коловратки также обладают, как и те, билатеральной нервной системой и мозгом, а также специализированными сенсорными и моторными нервами. Однако, мало отличаясь от инфузории размером, внешним видом и образом жизни, коловратки очень напоминают последних также поведением и не обнаруживают более высоких психических способностей, чем инфузории. Это опять показывает, что ведущим для развития психической деятельности является не общее строение, а конкретные условия жизнедеятельности животного, характер его взаимоотношений и взаимодействий с окружающей средой. Одновременно этот пример еще раз демонстрирует, с какой осторожностью надо подходить к оценке «высших» и «низших» признаков при сравнении организмов, занимающих различное филогенетическое положение, в частности при сопоставлении простейших и многоклеточных беспозвоночных.
Движения.
Кольчатые черви обитают в морях и пресноводных водоемах, но некоторые ведут и наземный образ жизни, передвигаясь ползком по субстрату или роясь в рыхлом грунте. Морские черви отчасти пассивно носятся течениями воды как составная часть планктона, но основная масса ведет придонный образ жизни в прибрежных зонах, где селится среди колоний других морских организмов или в расщелинах скал. Многие виды живут временно или постоянно в трубках, которые в первом случае периодически покидаются их обитателями, а затем вновь разыскиваются. Хищные виды отправляются из этих убежищ регулярно на «охоту». Трубки строятся из песчинок и других мелких частиц, которые скрепляются выделениями особых желез, чем достигается большая прочность построек. Неподвижно сидящие в трубках животные ловят свою добычу (мелкие организмы), подгоняя к себе и процеживая воду с помощью венчика щупалец, который высовывается из трубки, или же прогоняя сквозь нее поток воды (в этом случае трубка открыта на обоих концах).
В противоположность сидячим формам свободно-живущие черви активно разыскивают свою пищу, передвигаясь по морскому дну: хищные виды нападают на других червей, моллюсков, ракообразных и иных сравнительно крупных животных, которых хватают челюстями и проглатывают; растительноядные отрывают челюстями куски водорослей; другие черви (их большинство) ползают и роются в придонном иле, проглатывают его вместе с органическими остатками или собирают с поверхности дна мелкие живые и мертвые организмы.
Малощетинковые черви ползают и роются в мягком грунте или придонном иле, некоторые виды способны плавать. Во влажных тропических лесах некоторые малоще-тинковые кольчецы вползают даже на деревья. Основная масса малощетинковых червей питается детритом, всасывая слизистый ил или прогрызаясь сквозь почву. Но существуют и виды, поедающие мелкие организмы с поверхности грунта, процеживающие воду или отгрызающие куски растений. Несколько видов ведут хищный образ жизни и захватывают мелких водных животных, резко открывая ротовое отверстие. В результате добыча всасывается с потоком воды.
Пиявки хорошо плавают, производя туловищем волнообразные движения, ползают, роют ходы в мягком грунте, некоторые передвигаются по суше. Помимо кровососущих существуют также пиявки, которые нападают на водных беспозвоночных и проглатывают их целиком. Наземные пиявки подстерегают свои жертвы на суше (млекопитающих и людей), в траве или на ветках деревьев и кустарников (во влажных тропических лесах). Эти пиявки могут довольно быстро двигаться. В передвижении наземных пиявок по субстрату большую роль играют присоски: животное вытягивает сперва туловище, затем присасывается к субстрату головной присоской, притягивает к ней задний конец туловища (с одновременным сокращением последнего), присасывается задней присоской и т.д.
Итак, двигательная активность кольчатых червей как при локомоции, так и при добывании пищи отличается большим многообразием и достаточной сложностью. Обеспечивается это сильно развитой мускулатурой, представленной прежде всего так называемым кожно-мышечным мешком. Он состоит из двух слоев: внешнего (подкожного), состоящего из кольцевых волокон, и внутреннего, состоящего из мощных продольных мышц. Последние простираются, несмотря на сегментацию, от переднего до заднего конца туловища. Ритмичные сокращения продольной и кольцевой мускулатуры кожно-мышечного мешка обеспечивают локомоцию: червь ползет, вытягивая и сокращая, расширяя и сужая отдельные части своего тела. Так, у дождевого червя вытягивается (и сужается) передняя часть тела, затем то же самое происходит последовательно со следующими сегментами. В результате по телу червя пробегают «волны» сокращений и расслаблений мускулатуры.
У кольчатых червей впервые в эволюции животного мира появляются подлинные парные конечности: кольчецы (помимо пиявок) носят на каждом сегменте по паре выростов, служащих органами передвижения (кроме головного конца, где они служат ротовыми органами) и получивших название параподий. Параподии снабжены специальными мышцами, двигающими их вперед или назад. Зачастую параподий имеют ветвистое строение. Каждая ветвь снабжена опорной щетинкой и, кроме того, венчиком из щетинок, имеющих у разных видов различную форму. От параподий отходят и щупальцевидные органы тактильной и химической чувствительности. Особенно длинными и многочисленными последние являются на головном конце, где на спинной стороне располагаются глаза (одна или две пары), а в ротовой полости или на особом (выпячиваемом) хоботке — челюсти. В захвате пищевых объектов могут участвовать и нитевидные щупальца на головном конце червя.
У ряда многощетинковых и всех малощетинковых червей параподии редуцированы (отсюда и название последних), остались лишь посегментно расположенные пучки щетинок. Так, у дождевого червя на каждом сегменте находится по четыре пары очень коротких, неразличимых невооруженным глазом щетинок, которые, однако, наподобие параподии служат для передвижения животного: являясь достаточно крепкими подвижными рычагами, они обеспечивают вместе с сокращениями кожно-мышечного мешка поступательное движение червя. С другой стороны, растопыривая свои щетинки и упираясь ими в грунт, дождевой червь настолько прочно фиксирует свое тело в земле, что практически невозможно вытащить его оттуда в неповрежденном виде. У некоторых других малощетинковых червей щетинки развиты значительно сильнее и пред-
Органы чувств и сенсорные способности.
Большой интерес для познания психической деятельности низших многоклеточных беспозвоночных представляет устройство и функционирование их органов чувств, представленных также весьма различными образованиями в соответствии с общим уровнем организации животного.
У наиболее низкоорганизованных представителей беспозвоночных органы чувств еще очень слабо дифференцированы как в морфологическом, так и в функциональном отношении. У этих животных трудно выделить органы осязания, химической чувствительности и т.д. Очевидно, первичные органы чувств вообще были плю-ромодальными, т.е. они обладали лишь общей, присущей всей живой материи чувствительностью, но в повышенной степени. Существование таких плюромо-дальных чувствительных клеток является весьма вероятной гипотезой, но в настоящее время такие рецепторные клетки, очевидно, уже не существуют. Специализация таких клеток по отдельным видам энергии привела к появлению унимодальных рецепторных образований, которые, как правило, реагируют лишь на один специфический вид энергии. Так появились термо-, хемо-, механо-, фото- и другие рецепторы.
Дата добавления: 2016-08-07; просмотров: 577;