Часть III ЭВОЛЮЦИЯ ПСИХИКИ. 4 страница
Глава 2 ПЕРЦЕПТИВНАЯ ПСИХИКА.
Низший уровень развития перцептивной психики.
Перцептивная психика является высшей стадией развития психического отражения. Напомним, что эта стадия характеризуется, по Леонтьеву, изменением строения деятельности — выделением содержания деятельности, относящейся к условиям, в которых дан объект деятельности в среде (операции). По этой причине мы встречаемся здесь уже с подлинными навыками и восприятиями. Предметные компоненты среды отражаются уже как целостные единицы, в то время как при элементарной сенсорной психике имело место отражение лишь отдельных их свойств или суммы последних. Предметное восприятие обязательно предполагает определенную степень обобщения, появляются чувственные представления.
Разумеется, перцептивная психика, свойственная огромному числу животных, стоящих на разных ступенях эволюционного развития, обнаруживает в конкретных своих проявлениях большие различия. Поэтому необходимо и на этой стадии развития психики выделить отдельные уровни, из которых здесь будет рассмотрен низший.
На этом уровне находятся прежде всего высшие беспозвоночные — головоногие моллюски и членистоногие (по меньшей мере высшие). Характеристику рассматриваемого уровня психического развития мы дадим на примере насекомых — центрального класса типа членистоногих. (Кроме насекомых к членистоногим относятся ракообразные, паукообразные, многоножки и некоторые мелкие группы.) Насекомые - - самый многочисленный класс животных как по количеству видов, так и по количеству особей. Насекомые обитают повсюду на суше (во всех климатических зонах) — как на поверхности, так и в почве, во всех пресных водоемах, а также в воздухе, поднимаясь на высоту до двух километров.
Движения.
Огромное разнообразие «экологических ниш», занимаемых насекомыми и другими членистоногими, обусловило, естественно, развитие весьма различных форм приспособления в области строения и поведения этих животных. Наиболее существенным и характерным для эволюции членистоногих, особенно насекомых, было завоевание новой среды обитания — суши. Членистоногие — первые наземные животные в истории Земли. Переход на сушу был сопряжен с развитием особых органов передвижения — конечностей в виде сложных рычагов, состоящих из отдельных, соединенных суставами члеников, по которым членистоногие и получили свое название. Такие сложные рычаги (их аналогами являются конечности наземных позвоночных) наилучшим образом служат для передвижения по твердому субстрату путем отталкивания от него. Членистоногие в состоянии передвигаться по суше почти всеми возможными способами (ползание, ходьба, бег как по горизонтальным, так и по вертикальным поверхностям, прыгание, передвижение в толще грунта). В воде членистоногие плавают, ныряют, ползают и даже бегают по ее поверхности.
У насекомых, кроме того, в связи с переходом к жизни в воздушной среде впервые появился и совершенно новый способ передвижения — полет с помощью крыльев. Насекомые способны развить в полете очень большую скорость: у некоторых стрекоз она достигает 90 километров в час, у самца овода — даже 122 километра в час. Поистине поразительна частота взмахов крыльев у многих насекомых. Так, мотылек делает лишь 9 ударов в секунду, пчела же — 190, а комнатная муха — 330, но у некоторых мух и перепончатокрылых скорость вибрации крыльев достигает до тысячи ударов в секунду. Столь быстрые движения и соответственно частоты мышечных сокращений не наблюдаются ни у каких других животных.
Соответственно и крылевая мускулатура насекомых отличается исключительно сложным устройством: она состоит из множества различно располагающихся мышечных пучков, частично действующих как антагонисты. Большую сложность обнаруживают и другие отделы мышечной системы.
Вообще у насекомых (не считая крылевой мускулатуры насекомых) различают следующие отделы мышечной системы: мышцы туловища, заведующие его движениями: мышцы конечностей, с помощью которых движется вся конечность по отношению к туловищу; мышцы отдельных члеников конечностей, мышцы внутренних органов.
Как видно, мускулатура насекомых отличается высокой степенью дифференцированности и специализированности. К тому же подавляющее большинство их мышц -поперечнополосатые. Количество отдельных мускулов исключительно велико — полторы тысячи и более. Внушительна и сила мышц: насекомые способны таскать предметы, иногда в сто раз превышающие их собственный вес. В целом, как отмечал Догель, у членистоногих мышечная система беспозвоночных достигает «апогея своего развития» как в морфологическом, так и в физиологическом отношении.
Центральная нервная система.
В соответствии со сложной и высокодифференцированной организацией двигательного аппарата находится и сложное строение центральной нервной системы насекомых, которую, однако, мы можем здесь охарактеризовать лишь в самых общих чертах.
Как и у кольчатых червей, основу всей нервной системы составляют у насекомых метамерно расположенные ганглии, связанные друг с другом межсегментными нервными стволами. Эти стволы сближаются у большинства насекомых настолько, что образуют (хотя бы внешне) одну непарную нервную цепочку. Одновременно имеет место и слияние последовательных метамерных ганглиев.
В связи с отчетливо выраженной цефализацией в головном отделе слившиеся ганглии образовали мощный головной мозг в виде сложно устроенного надглоточного узла, в котором различают три отдела: протоцеребрум, дейтоцеребрум и тритоцеребрум. Передний отдел, протоцеребрум, развит сильнее остальных. Особое внимание заслуживают находящиеся в нем так называемые грибовидные (или стебельчатые) тела — ассоциативные и координирующие центры. В наибольшей степени они развиты у насекомых с особенно сложным поведением — у пчел, прежде всего рабочих особей, и муравьев. С протоцеребрумом связана и пара очень крупных и сложно устроенных зрительных долей, иннервирующих сложные глаза. Дейтоцеребрум (средний отдел головного мозга) иннервирует усики, тритоцеребрум иннервирует область рта и, кроме того, тесно связан с симпатической нервной системой. Вообще для насекомых характерна большая концентрация нервных элементов. Такая концентрация повышает возможности управления отдельными системами органов и поднимает общий морфофизиологический уровень организма как целостной единицы.
Нервная система, и в частности головной мозг, устроена у разных членистоногих, однако, весьма различно. Существенные различия обнаруживаются между низшими и высшими представителями даже в пределах классов, а иногда и отрядов. Так, например, отношение размеров мозга к объему туловища равняется у майского жука 1 : 3290, у муравья 1 : 280, а у пчелы 1 : 174.
На очень высоком уровне развития находится нервная система головоногих моллюсков, в корне отличаясь от таковой других моллюсков. Особенно это относится к их весьма сложно устроенному головному мозгу. Обращает на себя внимание исключительная концентрация мозговых ганглиев: ганглии слились воедино и образовали общую околопищеводную массу. Основания зрительных нервов образуют громадные вздутия -- оптические ганглии, которые своими размерами нередко превосходят сами церебральные узлы.
Нервы, отходящие от мозга к эффекторам, достигают очень большой мощности - до миллиметра и больше в диаметре, что в 5—10 раз превышает толщину нервов большинства позвоночных! Этим обеспечивается значительно большая скорость проведения нервных импульсов, чем у низших моллюсков. Соответственно огромной величине глаза исключительно развит и зрительный нерв. Выдающийся советский гистолог А.А.Заварзин показал большое сходство тонкого гистологического строения оптических центров головоногих и позвоночных.
Таким образом, мы видим, что глубокие различия между элементарной сенсорной и перцептивной психикой находят свое воплощение и в структуре нервной системы уже на низшем уровне перцептивной психики.
Сенсорные способности, таксисы.
Специфические условия, в которых живут насекомые, далеко идущее приспособление двигательной
активности при обилии и разнообразии качественно весьма различных агентов среды, управляющих их поведением, — все это обусловило появление у данных животных многих, и притом своеобразно устроенных, органов чувств. Большую роль в жизни насекомых играет зрение. В разном количестве у разных видов имеются простые одиночные глаза. Но главный орган зрения - - своеобразный сложный глаз, получивший название фасеточного, так как его поверхность состоит из отдельных, четко ограниченных участков — фасеток. Каждой фасетке соответствует самостоятельный глазок — омматидий, изолированный от своих соседей пигментной прокладкой и функционирующий независимо от них. Общий внешний покров глаза образует над омматидиями утолщения, обозначаемые как роговица или роговой хрусталик и играющие роль диоптрического аппарата глаза. Каждый омматидий содержит 7— 8 зрительных клеток, аксоны которых оканчиваются в оптических ганглиях мозга.
Благодаря такому устройству сложных глаз в мозг насекомого передаются «мозаичные» изображения воспринимаемых предметных компонентов среды. Конечно, это не означает, что насекомое действительно видит окружающий мир в виде мозаики. В мозгу поступающие из отдельных омматидиев зрительные раздражения синтезируются в целостную картину, как это имеет место и у позвоночных. Все же нам очень трудно представить себе, как на самом деле видит насекомое.
Еще в прошлом веке делались попытки установить путем фотографирования через фасеточные глаза четкость и дальность зрения насекомых и раков. Позже, в частности в 60-х годах нашего века, такие опыты повторялись, но едва ли возможно таким образом решить проблему, ибо, как известно, видит не глаз, а мозг. Насекомые, очевидно, весьма близоруки: ясное видение, например, бабочки-крапивницы возможно лишь на расстоянии приблизительно 2—3 сантиметров. От объектов, расположенных дальше (уже начиная с, 5—7 сантиметров), насекомые получают лишь расплывчатые изображения, но, очевидно, этого достаточно для их общей ориентировки в пространстве.
На расстоянии пчела способна увидеть лишь объекты в сто раз большей величины, чем это необходимо для зрения человека. Отдельный цветок она увидит лишь тогда, когда она приблизится к нему вплотную. Все эти особенности зрения насекомых вполне соответствуют тому микроландшафту, в котором живут эти животные.
Острота зрения (разрешающая способность, т.е. способность видеть две сближенные точки) зависит у насекомых при прочих равных условиях, в частности, от количества омматидиев, которое, однако, сильно колеблется даже в пределах отрядов. Так, в отряде жуков это количество колеблется от трех-четырех десятков до нескольких десятков тысяч. У пчелы каждый глаз состоит приблизительно из четырех тысяч фасеток. Чем мельче фасетки, тем слабее в них освещение, но тем детальнее зрение. Особенно большое значение это имеет для предметного зрения. Кроме того, при более выпуклом глазе оси омматидиев сильнее расходятся, и в результате расширяется поле зрения. Одновременно суживается точка, видимая одной фасеткой, чем также увеличивается острота зрения.
Наилучшим образом насекомые видят подвижные объекты или неподвижные предметы во время собственного движения. Пролетающая над лугом пчела, например, видит покачивающиеся на ветру цветы или же изображение неподвижного цветка скользит по ее фасеточному глазу. В любом случае это подвижное (расплывчатое) изображение заставляет ее спуститься вниз и обследовать растение с минимального, т.е. оптимального для зрения пчелы, расстояния.
Что касается цветового зрения, то видимый ими спектр шире, чем у человека, так как его граница проходит в ультрафиолетовом секторе. Спорный вопрос о способности насекомых к восприятию красного цвета в настоящее время, очевидно, решен в положительном смысле (за исключением медоносной пчелы и некоторых других насекомых). Советский энтомолог Г.А.Мазохин-Поршняков считает, что вообще цветовое зрение у взрослых насекомых представляет универсальное явление. По его данным, цвета различаются даже видами, активная жизнь которых протекает ночью.
До сих пор мы касались только реакции насекомых на свет, который играет в их жизни очень большую роль. Не меньшее, если не большее значение имеют для них запахи. В естественном поведении насекомых стимулы различной модальности всегда выступают в комплексе. Так, уже упомянутые осы рода Philanthus охотятся за пчелами, руководствуясь зрением с расстояния 30 сантиметров, но опознают их по запаху лишь с расстояния 15—5 сантиметров. Во время зрительного поиска оса совершенно не реагирует на запах пчелы.
Аналогично обстоит дело и у других членистоногих. Водяные пауки рода Pirata ориентируются прежде всего по вибрации поверхности воды, в непосредственной же близости от жертвы — зрением (рис. 36). Другие пауки узнают о попадании в их сеть насекомого, воспринимая вибрации паутины. Но обследование жертвы производится сразу тремя способами: паук прикасается к ней передними конечностями, снабженными хеморецепторами, а также так называемыми щупиками (органы осязания), и, кроме того, паук на вкус пробует жертву укусом.
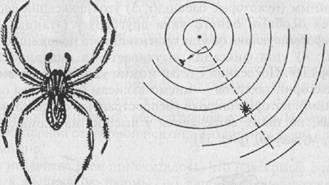
Рис. 36. Водяной паук Pirata бежит по поверхности воды к источнику вибрации (центральная точка). Только непосредственно около неподвижной мухи вид последней приобретает преобладающее влияние и паук сворачивает к ней (опыт Берестынской-Вильчек)
Для ознакомления с таксисами у насекомых рассмотрим в качестве примера фототаксисы. Эти таксисы (положительные или отрицательные) встречаются у насекомых прежде всего в виде тропо-, тело- и менотаксисов, хотя не утратили своего значения и примитивные таксисы. Так, например, положительные и отрицательные клинотаксисы с четко выраженными маятникообразными поисковыми движениями играют ведущую роль в оптической ориентации у многих личинок мух. Тропо- и телотаксисы связаны у них друг с другом многими переходными формами. У взрослых насекомых чаще всего встречаются, однако, четко выраженные телотаксисы. Как правило, тропотаксисное движение очень скоро переходит у них в телотаксисное. У комаров и вшей преобладает тропотак-сисная ориентация движения.
Менотаксисы, относящиеся к более сложным формам ориентации, встречаются у высших насекомых (муравьев, пчел, бабочек, жуков). Немецкий исследователь поведения насекомых М.Линдауер выделяет здесь следующие четыре случая: 1) угол фиксации источника света случаен (муравьи, жук-навозник); 2) угол фиксации является врожденным (некоторые бабочки); 3) угол фиксации сообщается особями одного вида друг другу («танцы» пчел, информирующие об угле ориентации по положению солнца); 4) угол фиксации выучивается в индивидуальном порядке. Последний случай можно уже отнести к высшей категории таксисов — мнемотаксисам (выученная ориентация), которые широко распространены среди позвоночных, но также встречаются у высших насекомых (пчел, ос, муравьев) (рис. 37).

Рис. 37. Мнемотаксис у роющей осы Philanthus. После отлета осы кольцо из шишек вокруг норки было перемещено в сторону. После возвращения оса ищет норку внутри кольца и находит ее лишь после повторных облетов (опыт Тинбергена)
Необходимо, однако, подчеркнуть, что и в тех случаях, когда ориентация осуществляется на основе других фототаксисов, поведение насекомых одновременно определяется рядом дополнительных факторов. Например, доказано, что у многих насекомых характер их фототаксисного поведения сильно видоизменяется в зависимости от температуры воздуха, времени дня, физиологического состояния и т.д. Хорошо известно, что муха не реагирует на свет, пока спокойно летает по комнате. Но стоит начать ее преследовать, как у нее сейчас же «просыпается» положительный фототелотаксис: она быстро направляется к источнику света, к окну.
Аналогичным образом насекомые реагируют на разнообразные механические, температурные, химические и оптические раздражения, на силу тяжести, и соответственно у них встречаются во многих различных формах тигмо-, рео-, анемо-, гидро-, гео-, термо-, хемо-, фото- и другие таксисы.
Предметное восприятие.
Особый интерес представляет способность насекомых (и других членистоногих) к оптическому восприятию форм как необходимого компонента перцептивной психики. На уровне элементарной сенсорной психики различение форм еще невозможно.
До недавнего времени считалось, что насекомые способны к восприятию формы, но лишь в специфических рамках. Эта специфика усматривалась в том, что в экспериментах насекомые, особенно пчелы, оказывались способными различать лишь фигуры, напоминающие по своим очертаниям цветы (например, кресты или звездчатые фигуры), простые же геометрические фигуры они в этих опытах не различали. Однако Мазохин-Поршняков (в работах, проведенных совместно с Г.М.Вишневской) показал, что пчел можно с полным успехом дрессировать и на такие фигуры, как треугольник, круг, квадрат, из чего он заключил, что пчелы способны распознавать фигуры непосредственно по их графическим признакам.
О способности насекомых к распознаванию форм свидетельствуют и полевые опыты Тинбергена, в которых самки роющих ос обучались распознавать круг из сосновых шишек, выложенных вокруг входа в норку. После того как оса улетала за добычей, круг передвигался на 30 сантиметров в сторону. После возвращения оса искала норку сперва в центре круга (рис. 37). В следующих опытах к тому же шишки заменялись черными камешками, а вокруг норки выстраивался треугольник или даже эллипс из шишек, но оса тем не менее прилетала в круг, хотя из предыдущих опытов было известно, что оса вполне способна отличать камешки от шишек. Таким образом, пространственная ориентация осуществлялась здесь только по форме (кругу).
Хотя у высших насекомых способность к предметному восприятию и не вызывает сомнения, в этом отношении они явно уступают позвоночным. Очевидно, это связано со специфическим строением органов зрения: разрешающая способность глаза зависит у насекомых от количества омматидиев. Чем их больше, тем выше эта способность, так как картина воспринимаемого объекта расчленяется на большее число точек. К тому же насекомые, как и другие членистоногие, очевидно, ориентируются преимущественно все же не по предметным компонентам среды как таковым, а по отдельным их признакам, как это имеет место на стадии элементарной сенсорной психики.
У других беспозвоночных, находящихся также на низшем уровне стадии перцептивной психики, — у головоногих моллюсков - - зрение играет не меньшую, если не большую роль, чем у насекомых, и является у них, безусловно, ведущей рецепцией (хотя эти животные ориентируются и хорошо развитыми осязанием и химической чувствительностью). Соответственно и орган зрения получил у головоногих исключительное развитие: головоногие обладают наиболее крупными глазами не только среди обитателей морей, но и вообще в животном мире, причем не только в относительном, но и в абсолютном отношении. Глаза гигантских спрутов (род Architeuthus) достигают в поперечнике 40 сантиметров при общей длине животного (считая и «руки») до 20 метров. Для сравнения можно указать, что у всех, даже самых крупных, китов (например, у 30-метрового голубого кита) диаметр глаза составляет немногим больше десяти сантиметров. Поразительна и огромная разрешающая способность (зоркость) глаза головоногих: на один квадратный миллиметр приходится у разных представителей головоногих от 40 000 до 162 000 палочек, у человека — 120 000 — 400 000, у совы, обладающей самым зорким глазом в мире, — 680 000. Что же касается предметного восприятия, то головоногие моллюски в отличие от низших моллюсков уже способны к подлинному предметному восприятию, что выражается прежде всего в различении ими формы объектов. Так, осьминог в состоянии отличить треугольник от квадрата, малый квадрат от большого, крест от квадрата, вертикальный прямоугольник от горизонтального, треугольник от ромба и т.д. Всего, как удалось установить, осьминог способен различать 46 разных форм (опыты Б.Б.Бойкотта и Дж.З.Янга). Все же зрение головоногих, хотя и сближается с таковым позвоночных, отличается и рядом примитивных признаков, делающих отражение предметов действительности недостаточно полноценным. Еще предстоит большая исследовательская работа для выяснения специфических особенностей оптического восприятия головоногих.
Общение и групповое поведение.
У высших беспозвоночных уже вполном объеме представлена категория поведения, которая, как было показано, только в зачаточной форме обнаруживается у некоторых представителей низших многоклеточных беспозвоночных. Речь идет об общении, которое у высших беспозвоночных проявляется иногда и в очень сложных формах подлинного группового поведения (в отличие от колониальной жизни, свойственной многим низшим беспозвоночным). Напомним, что общение, в точном значении этого термина, появляется только тогда, когда контакты между животными начинают осуществляться посредством специальных сигнальных действий.
У головоногих моллюсков и членистоногих, особенно у насекомых, мы находим четкие системы коммуникации с передачей информации по разным сенсорным каналам. Это относится прежде всего к пчелам и муравьям, что, конечно, связано со сложной, высокодифференцированной структурой их сообществ и разделением функций между членами последних (распределение пищедобывательной, защитной и воспроизводительной функций между отдельными особями).
У муравьев разные формы химической чувствительности играют, очевидно, большую роль, чем другие виды рецепции, и общение между особями осуществляется прежде всего по этому каналу: личинки муравьев выделяют специфические вещества, побуждающие взрослых рабочих особей кормить их, по запаху обитатели одного муравейника узнают «своих» и «чужих». Даже о том; жив или мертв муравей, его сородичи узнают по специфическим выделениям.
Согласование поведения обитателей муравейника происходит путем подачи и приема химических сигналов. Достаточно указать на химические средства муравьев (а также пчел и ос), выделение которых служит сигналом тревоги. Эти вещества выделяются муравьями в случае опасности и быстро распространяются по воздуху. Их восприятие вызывает у других муравьев («солдат») защитную реакцию (готовность к нападению на врага), хотя источник опасности им еще не известен. Поскольку, однако, радиус действия такого сигнала, подаваемого одним муравьем (например, в случае вторжения в муравейник какого-либо насекомого), невелик, то он способен указать и место, откуда угрожает опасность. Чем больше эта опасность, тем больше муравьев выделяют химические сигналы тревоги и тем больше, следовательно, особей призывается на помощь.
По другим выделениям, оставляемым на субстрате в виде следа, муравьи узнают, куда, в какую сторону ушел их сородич. К тому же интенсивность такого пахучего следа указывает на количество пищи, обнаруженной в том месте, куда ведет след, тем более что, направляясь к муравейнику, муравьи наносили по пути пахучие метки лишь в тех случаях, когда они находили корм. При этом тем больше оставляется меток, чем больше обнаружено корма. Поскольку эти метки довольно быстро испаряются, вновь идущие по этому следу муравьи будут воспринимать лишь свежие метки, а таких будет все меньше по мере приближения кормового объекта. Таким образом, муравьи в конце концов узнают о том, что пищевые ресурсы в данном месте исчерпаны, и перестанут к нему направляться.
У некоторых пчел также существует химический способ передачи информации о нахождении пищи. Это относится, прежде всего, к пчелам - из подсемейства Meliponinae, у которых нет «языка танца». Эти широко распространенные в тропиках пчелы не имеют жала, но яростно кусаются. У мелких форм (самая маленькая имеет длину всего 1,5 миллиметра) нашедшая пищу сборщица только приводит других пчел в возбужденное состояние, беспокойно бегая по сотам, толкая их и время от времени подбегая к летку. Вылетающие же пчелы ориентируются только по запаху цветов. У других видов возбуждение пчел первоначально достигается подобным же образом, но дополнительно сборщица, принесшая взяток, приводит соты толчками в вибрирующее состояние. После этого она возвращается к обнаруженным цветам, а на обратном пути оставляет на траве, ветках или камнях через каждые два-три метра пахучие метки, по которым затем приводит группами других пчел к источнику пищи. Это достигается тем, что пчела-разведчица летает взад и вперед между двумя соседними метками, пока другие пчелы не присоединяются к ней. Следовательно, здесь мы имеем дело с комплексной передачей информации по оптическому, тактильно-вибрационному и, возможно, акустическому каналам, но при этом ведущую роль играют оставляемые сборщицей запахи.
У нашей медоносной пчелы также существует химический способ передачи информации, но только как вспомогательный, когда она имеет дело с кормом, лишенным собственного заметного запаха. В этих случаях место нахождения кормового объекта метится секретом из так называемой железы Насонова.
Однако основным и наиболее совершенным способом передачи информации о кормовых объектах являются «танцы». Найдя источник пищи (нектара) и вернувшись в улей, пчела раздает другим пчелам-сборщицам пробы нектара и приступает к «танцу», который состоит из пробежек по сотам. Рисунок танца зависит от местонахождения обнаруженного источника пищи: если он находится рядом с ульем (на расстоянии 2-5 метров от него), то производится «танец-толчок»: пчела беспорядочно бежит по сотам, время от времени виляя брюшком; если же корм обнаружен на расстоянии до 100 метров от улья, то выполняется один из описанных К.Фришем танцев — «круговой», который состоит из пробежек по кругу попеременно по часовой стрелке и против нее (рис. 38). Если же нектар обнаружен на большем расстоянии, то выполняется «виляющий» танец, также описанный и изученный Фришем. Это пробежки по прямой, сопровождающиеся виляющими движениями брюшка с возвращением к исходной точке то слева, то справа (рис. 38, а). Интенсивность виляющих движений указывает на расстояние находки: чем ближе находится кормовой объект, тем интенсивнее выполняется танец (рис. 38, б).
Кроме того, как тоже показал Фриш, во второй форме танца угол между линией пробежки и вертикалью (на вертикально расположенных сотах) соответствует углу между линией полета пчелы от улья к кормовому объекту и положением солнца. Таким образом, горизонтальное направление истинного полета переносится на вертикальную плоскость, ориентация по положению солнца подменяется ориентацией по земному притяжению. При этом, если следует лететь навстречу солнцу, пчела танцует снизу вверх, если же от солнца, то сверху вниз (рис. 38, в). Таким образом, медоносная пчела способна к транспозиции с фотоменотаксисов на геоменотаксисы: фиксированный угол к видимому положению солнца переносится в темном улье к силе тяжести.
Эта способность медоносной пчелы к транспозиции, коренящаяся в широко распространенной среди насекомых способности к переносу менотаксисных движений на вертикальную, плоскость, обеспечивает; как мы видим, исключительную полноту и точность передачи информации. Такая транспозиция является частным случаем трансформации, лежащей в основе всякого общения между животными: свойства действительных предметов и ситуации трансформируются на языке животных с помощью видоспецифического «кода» в оптические, акустические и прочие условные сигналы, информирующие других осо  бей об этих свойствах. Несмотря на всю сложность и точность, система коммуникации медоносной пчелы в принципе не отличается от любой другой системы общения у животных. Переводятся ли истинные пространственные параметры в сигналы, воспринимаемые с помощью зрения, осязания и чувства силы тяжести, как это имеет место у медоносной пчелы, или же в сигналы, воспринимаемые хеморецепцией (муравьи), -- информация в любом случае передается в трансформированном, условном виде. Здесь как нельзя нагляднее выступают характер и функциональное значение психики у животных.
бей об этих свойствах. Несмотря на всю сложность и точность, система коммуникации медоносной пчелы в принципе не отличается от любой другой системы общения у животных. Переводятся ли истинные пространственные параметры в сигналы, воспринимаемые с помощью зрения, осязания и чувства силы тяжести, как это имеет место у медоносной пчелы, или же в сигналы, воспринимаемые хеморецепцией (муравьи), -- информация в любом случае передается в трансформированном, условном виде. Здесь как нельзя нагляднее выступают характер и функциональное значение психики у животных.
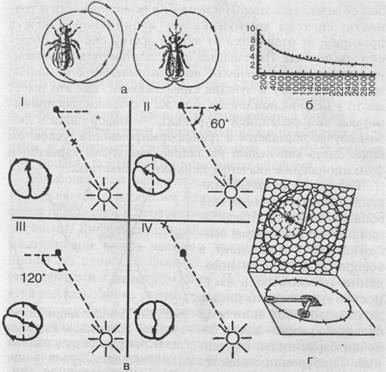
Рис. 38. «Танцы пчел». Обозначения: а — «круговой танец» (слева) и «виляющий» (справа); б — интенсивность выполнения «виляющих танцев» как показатель расстояния до местонахождения источника пищи (на оси ординат отмечено число «танцев» за 15 секунд, на оси абсцисс — расстояние до пищи в метрах); в — «виляющий танец» как показатель направления полета к источнику пищи: I — навстречу солнцу, II — под углом 60° налево от положения солнца, III — направо от положения солнца, IV — от солнца. Точка — местонахождение улья, крестик — местонахождение пищи. Изображение «танцев» следует представить себе на вертикальной плоскости (по Фришу); г — схема, иллюстрирующая транспозицию угла полета по отношению к положению солнца (горизонтальная плоскость) на вертикальную плоскость сот, на которых пчела выполняет «виляющий танец» (по Кэстнеру)
Вершиной развития инстинктивных компонентов общения является ритуализа-ция поведения, которая особенно четко выступает в воспроизводительном поведении насекомых, в частности в «ухаживании» самцов за самками. Вот типичный брачный «ритуал» самца ба-бочки-бархатницы, описанный Тинбергеном: полетев вслед за самкой, самец садится рядом с ней, потом становится перед ней, начинает «дрожать» крыльями (передне-задние движения сомкнутых крыльев), затем повторно раскрывает и закрывает их, что сопровождается постукиванием усиками по крыльям самки. После этого самец обхватывает самку крыльями и зажимает между ними ее усики, которые при этом прикасаются к пахучему участку на крыльях самца. Только после этого, если самка не отвергает «ухаживания» самца, начинается спаривание.
Дата добавления: 2016-08-07; просмотров: 594;