Гибридологический анализ количественных признаков
Допустим, мы имеем дело с двумя изогенными линиями Р1 и Р2, принадлежащими одному виду. Несмотря на одинаковые условия среды, их средние значения могут сильно различаться, особенно если линии выделены из географически удаленных популяций. Мы исходим из того, что число генов в геномах сравниваемых линий одинаково, поэтому наследственные различия между ними вызваны разным набором аллелей одних и тех же локусов. Представим себе чисто условно, что в генотипе линии Р1 (с маленьким значением признака) собраны «слабые» аллели, которые будем обозначать строчными буквами, а в генотипе линии Р2 (с большим значением признака) – «сильные» аллели, для их обозначения будем применять прописные буквы.
Пусть разница средних значений признака двух изогенных линий обусловлена отличием в силе аллелей только одного локуса. Тогда генотип линии Р1 обозначим аа, а генотип линии Р2 – АА. Примем еще одно упрощающее условие: пусть по степени доминирования оба аллеля (а и А) равны (h = 0,5). Теперь введем представление об эффекте аллельного замещения. Будем считать, что замещение одного слабого аллеля на сильный увеличивает генотипическое значение признака на α единиц. Тогда замещение обоих слабых аллелей на сильные увеличит это значение на 2α единиц. Вспомним, что генотипическое значение признака равно его среднему значению у особей с одинаковым генотипом, т. е.
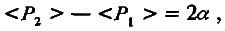
(4.6)
символ < > означает среднее значение признака в линии.
Проведя массовые скрещивания особей таких линий, получим популяцию гибридов F1 :
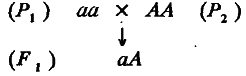
Все особи популяции F1 имеют один и тот же генотип аА, поэтому изменчивость признака в данном случае обусловлена исключительно средой. Будем считать, что средовая дисперсия для всех популяций (F1, P1 и Р2 ) одинакова и равна σe2. Среднее значение признака у особей F1 (обозначим его т ) должно равняться генотипическому значению гетерозигот аА, т. е. оно должно на α единиц превосходить среднее значение особей линии P1 и настолько же уступать среднему значению особей Р2. Иными словами, m попадает точно в середину интервала между средними значениями признака обеих родительских популяций. Тогда средние значения признака всех рассмотренных популяций можно представить как
<F1> = m ; <Р1 > = m – а; <Р2> = m + а . (4.7)
Итак, m, m – а и m + а – генотипические значения признака у особей с генетической конституцией аА, аа и АА соответственно.
Перейдем к популяции F2 , возникшей или при самооплодотворении, или при панмиктическом скрещивании особей из F1 :
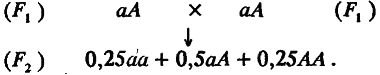
Из этой схемы видно, что вся совокупность особей F2 разбивается на три генотипических класса; каждый из них можно охарактеризовать его долей в выборке и средним значением признака. Одна четверть особей имеет генотип аа, другая четверть – генотип АА и половина – генотип аА. Поскольку средние значения признака у особей с этими генотипами равны соответственно m – а, m + а и m, то по (4.3) и (4.4) легко рассчитать среднее значение (М) и дисперсию (σg2) для распределения особей по генотипическим классам:
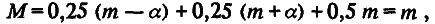
(4.8)
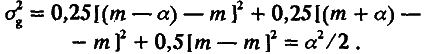
(4.9)
Таким образом, популяция F2 обладает дисперсией (а2 /2), обусловленной различием особей по генотипу. Кроме того, из‑за «шума» среды популяция обладает и средовой дисперсией σe2. Этот шум не сдвигает средних значений, поэтому <F2> = m. Эффекты среды и генотипа независимы, отсюда следует, что дисперсия по признаку в поколении F2 должна быть больше средовой на положительную величину а2 /2, т. е.
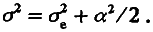
(4.10)
Теперь попробуем рассмотреть более общий случай, когда особи двух изогенных линий различаются аллелями n несцепленных локусов. По‑прежнему будем считать, что все слабые аллели собраны у линии P1, а все сильные – у линии Р2. Проведем их скрещивание:
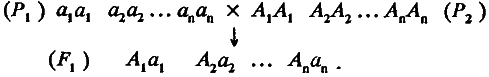
При оценке среднего значения популяции F1 сделаем два предположения: во‑первых, по степени доминирования все аллели равны и, во‑вторых, замещение в каждом локусе одного слабого аллеля на сильный увеличивает генотипическое значение признака на одну и ту же величину а. Следовательно, разность средних значений родительских популяций должна быть равна 2na, а среднее значение признака в популяции F, (обозначим его <F1> ) будет находиться в точке m, т. е. точно посередине между средними значениями родительских линий. Такая модель, где вклады всех аллелей в величину признака суммируются, получила название аддитивной. Главным основанием для ее применения является попадание среднего значения признака в популяциях F1 и F2 посередине между средними значениями родительских популяций. Итак, для случая n локусов
<F1 > = m; <P1> = m – na; <Р2> = m + na . (4.11)
Так как генотип всех особей F1 одинаков, то изменчивость признака в этой популяции обусловлена только влиянием среды, и ее дисперсия равна σe2.
Теперь перейдем к популяции F2, представляющей собой смесь огромного числа (3n) генотипов. Формулу генотипа каждой особи можно записать как ряд из n аллельных пар со случайной комбинацией сильных и слабых аллелей в каждой паре. Поскольку аллельный состав каждого локуса формируется независимо от остальных, то генотипическая дисперсия популяции F2 должна представлять собой сумму дисперсий, каждая из которых отражает варьирование у разных особей числа сильных аллелей в каком‑то одном локусе. Напомним, что в данной, аддитивной, модели замещение в любом локусе слабого аллеля на сильный ведет к увеличению генотипического значения признака на одну и ту же величину а. Отсюда следует (см. (4.9)), что каждый из n локусов вносит в генотипическую дисперсию поколения F2 один и тот же вклад а2 /2. Итак, величину фенотипической дисперсии σ2 в популяции F2 можно передать формулой
σ2 = σe2 + na2 /2. (4.12)
Это равенство вместе с другим
<Р2> – <Р1 > = 2na (4.13)
образует систему двух независимых уравнений, позволяющих определить величину п:
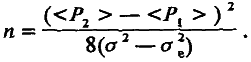
(4.14)
Хотя эта знаменитая формула Кастла – Райта верна лишь в рамках аддитивной модели, она дает возможность ориентировочно подойти к числу генетических факторов, ответственных за межлинейную разницу величины признака.
Что нам дал этот гибридологический экскурс? Очень много. Хотя природные популяции – это не поколение F2 но и здесь генотипическое значение признака можно считать суммой n независимо варьирующих слагаемых, где n – число локусов в генофонде популяции. Только в отличие от F2 число аллелей каждого локуса в данном случае может быть больше двух, и в пары они соединяются не в отношении 1:2:1, а по закону Харди – Вайнберга. Хотя мы ничего не знаем ни об эффектах этих аллелей, ни о степени их доминирования, ясно одно: популяционная дисперсия признака должна расти с увеличением числа локусов, принимающих участие в его формировании.
Сигма
Очень часто в качестве меры фенотипической изменчивости используют квадратный корень из дисперсии – так называемое среднеквадратичное отклонение (σ). Для экономии места будем именовать эту величину сигмой , по названию греческой буквы, обычно используемой для ее обозначения. Измеряемая в единицах величины самого признака, сигма очень удобна как масштаб для оценки отклонения величины признака от среднепопуляционного значения. Если признак имеет нормальное распределение, то доля особей с отклонением в пределах одной сигмы составляет 68 %, двух сигм – 95 и трех сигм – 99,7 %. В связи с этим полный размах изменчивости признака, распределенного по нормальному закону, попадает в интервал ±3σ (закон трех сигм). В сигмах принято измерять разность средних значений сравниваемых распределений и, в частности, эффект аллельных замещений.
Как и дисперсия, сигма может служить мерой средовой изменчивости – шума среды. Мы видели, что формула Кастла – Райта позволяет приближенно оценить число локусов, ответственных за различие линий по величине количественного признака. Зная это число, можно определить и средний эффект замещений слабого аллеля на сильный. Очень часто эффект оказывается меньше величины средовой сигмы σe. Это обстоятельство позволяет почувствовать основную трудность, с которой сталкивается исследователь генетических основ количественных признаков. Ведь если эффект аллельных замещений меньше шума среды, то по фенотипу особи определить ее генотип невозможно. Это же объясняет и гладкий характер распределений по количественным признакам. Шум среды как бы замазывает генотипические различия особей.
При анализе природных популяций широко используется еще одна мера – коэффициент изменчивости признака, т. е. отношение сигмы к среднему значению, выраженное в процентах. Наиболее поразительная черта этого коэффициента заключается в его стабильности при переходе от популяции к популяции в пределах одного вида и даже при сравнении популяций разных видов одного рода. Более того, коэффициент изменчивости самых разных признаков в популяциях эволюционно далеких видов также не слишком различается, несмотря на большие расхождения в величинах средних значений. Отсюда следует, что между сигмой и средним значением существует связь, близкая к прямой пропорциональности. Однако такой вывод в корне противоречит нашему исходному представлению о нормальном характере распределения особей по признаку.
Напомним, что нормальное распределение однозначно определяется двумя независимыми параметрами – средним значением и дисперсией. Иными словами, если признак распределен в каждой популяции по нормальному закону, то при переходе от одной популяции к другой сигма и среднее значение должны изменяться независимо. Выходит, что наша первоначальная интерпретация средового влияния как суммарного действия большого числа легких независимых толчков не совсем точна. Правда, уже со времен создателя биометрии Ф. Гальтона известно, что при измерении признака логарифмической шкалой связь между сигмой и средним значением зачастую теряется. Если обозначить сдвиг значения признака X, измеренного обычной (арифметической) шкалой, как ΔХ, то отношение этого сдвига к величине признака (его относительный сдвиг) примерно равно изменению признака, измеренного в логарифмическом масштабе, поскольку
ΔX / X ≈ ΔlnX. (4.15)
Распределение, которое становится нормальным после логарифмирования значения случайной величины, называется логарифмически‑нормальным. Его характерной особенностью является линейная связь между сигмой и средним значением. Вообще говоря, логарифмически‑нормальное распределение совсем не симметрично и обладает длинным «хвостом», плавно спускающимся в сторону увеличения признака. Однако так оно выглядит при большом размахе изменчивости признака в арифметической шкале (например, в 10 раз). Если же такой диапазон значительно скромнее (скажем, только в 1,5 раза), то распределение по признаку в обеих шкалах выглядит очень сходным, и даже в арифметической шкале не слишком отличается от нормального. Поэтому при анализе особей одной популяции, когда размах изменчивости невелик, логарифмически‑нормальные распределения выглядят как нормальные.
Опыт биометрии свидетельствует, что наследование количественных признаков, измеренных с помощью логарифмической шкалы, часто удовлетворяет аддитивной модели. Это означает, что межлинейное различие по величине логарифма признака можно трактовать как сумму эффектов аллельных замещений по ряду локусов. Заметим, что логарифм числа представим в виде суммы близких по величине слагаемых, когда само это число является произведением близких по значению сомножителей. Выходит, что замещение слабого аллеля на сильный в локусе, ответственном за развитие количественного признака, ведет к увеличению его генотипического значения в какое‑то число раз. Причем это число не слишком различается при аллельных замещениях в разных локусах.
Дата добавления: 2016-01-26; просмотров: 1030;