Негативные эффекты искусственного отбора
Первый лабораторный эксперимент по отбору на изменение количественных признаков был проведен в начале XX века американским генетиком У. Кастлом. Он ставил перед собой цель – превратить пегих (пятнистых) крыс в одноцветных. В одном варианте отбор был направлен на увеличение площади белых пятен, в другом – в противоположном направлении. Примерно через 20 поколений Кастл был близок к цели, т. е. ему удалось получить почти белых и почти черных крыс, но опыт пришлось прекратить, так как животные захирели и перестали размножаться. Короче говоря, у крыс резко снизились основные показатели приспособленности.
В дальнейшем сходные результаты были получены на большом числе животных объектов – мышах, курах, золотых рыбках, мучных хрущаках и, конечно, на дрозофиле. Везде по мере сдвига самых разных количественных признаков в любом направлении наблюдалось снижение показателей приспособленности: прежде всего жизнеспособности и плодовитости. Популяции как бы сопротивлялись действию искусственного отбора.
Особенно много экспериментов проведено по увеличению числа щетинок на разных частях тела у дрозофилы. Пристрастие именно к этому объекту объясняется очень коротким жизненным циклом плодовой мушки и ее хорошей генетической изученностью. Пример такой селекции приведен на рис. 14. Величина признака сначала монотонно возрастает, затем (обычно после 15–20 поколений отбора) наступает застой, ответ на отбор прекращается, а жизнеспособность и плодовитость мух падают. Оказывается, если отбор прекратить, то основные показатели приспособленности довольно быстро восстанавливаются, однако обычно это сочетается с существенным снижением числа щетинок, т. е. за несколько поколений «отдыха» от отбора величина признака сдвигается назад в сторону его исходных значений. Если после периода «отдыха» отбор возобновить, то можно снова поднять признак на прежний уровень, иногда даже при более высоких значениях главных компонент приспособленности.
Для экспериментов по долговременному отбору на увеличение числа щетинок характерна поразительная невоспроизводимость динамики изменения признака. В то же время в этой картине при всем ее многообразии имеются некоторые общие моменты. Обычно все начинается с довольно быстрого ответа на отбор, потом скорость увеличения признака постепенно падает, и кривая отбора выходит на плато. В этом состоянии «застоя», несмотря на продолжающуюся селекцию, популяция может находиться неопределенно долго. Затем внезапно может наступить фаза быстрого ответа на отбор, и кривая выходит на новое, более высокое плато. Этап нового застоя через какое‑то число поколений может опять смениться фазой внезапного ответа, потом снова наступает застой и т. д.
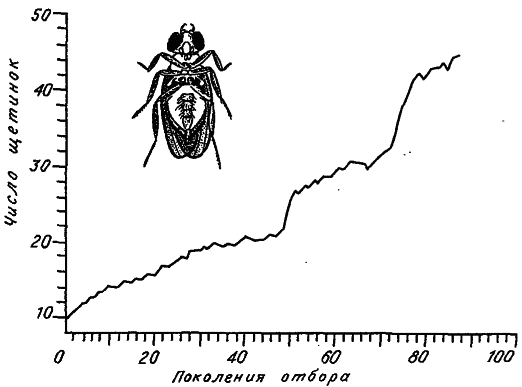
Рис. 14. Отбор на увеличение числа абдоминальных щетинок у дрозофилы Drosophila melanogaster (по: [Yoo, 1980]).
Вся эта феноменология выглядит весьма загадочно и до сих пор не получила удовлетворительного объяснения. Существует несколько гипотез. Самая простая из них связывает падение приспособленности с так называемой инбредной депрессией. Действительно, обычно численность лабораторных понуляций невелика и в каждом поколении большая их часть подвергается браковке. Поэтому с ходом отбора постепенно повышается степень родства особей, вступающих в скрещивание. А это означает прогрессивное возрастание степени гомозиготности генов, часть которых может быть представлена вредными рецессивными аллелями. Однако расчеты показывают, что инбредная депрессия повинна лишь в части негативных последствий селекции.
Вторая гипотеза, высказанная выдающимся американским генетиком и математиком С. Райтом, объясняет негативный эффект селекции нарушением взаимодействия генов, хорошо «подогнанных» друг к другу естественным отбором. В ходе искусственного отбора в генофонде популяции нарастает концентрация аллелей, которые сдвигают признак в направлении, нужном экспериментатору, но плохо «сочетаются» с аллелями других генов.
Третья гипотеза принадлежит знаменитому английскому генетику К. Мазеру. Он предположил, что степень развития признака определяется совокупным эффектом большого числа локусов, аллели которых могут сдвигать величину признака в любом направлении. По мнению Мазера, слабые и сильные аллели (по эффекту на признак) разбросаны вдоль хромосомы случайно, а между ними находятся гены, влияющие на приспособленность и которые также представлены своими сильными и слабыми аллелями.
При отборе, направленном на увеличение признака, сначала отбираются особи – носители хромосом с повышенным содержанием сильных аллелей. Затем в результате кроссоверной рекомбинации (т. е. обмена участками между гомологичными хромосомами) синтезируются хромосомы с еще большим числом сильных аллелей, однако новые (рекомбинантные) хромосомы могут нести увеличенное число аллелей, снижающих приспособленность. В итоге возникают генотипы, удовлетворяющие экспериментатора, но неудовлетворительные с точки зрения естественного отбора. Гипотеза неплохо объясняет длительные периоды «застоя» на кривых отбора как время ожидания синтеза рекомбинантных хромосом.
Четвертая группа гипотез объясняет картину отбора мутациями. В связи с этим мы должны сначала кратко ознакомиться с основными открытиями Мукаи. В начале 60‑х годов этот японский генетик приступил к серии экспериментов, продолжающихся и по сей день. На начальном этапе своих исследований Мукаи попытался оценить интенсивность и характер мутационного давления на гены, ответственные за жизнеспособность мух в лабораторных условиях.
Для этого была заложена серия линий, причем геном каждой из них содержал копию одного вполне конкретного варианта второй хромосомы. В каждом поколении из хромосомного фонда линии случайно извлекался один экземпляр второй хромосомы и размножался. Заметим, что выбираемая хромосома передавалась от поколения к поколению без оценки ее влияния на приспособленность, т. е. без отбора. Кроме того, генетическая схема опыта позволяла на любом поколении сопоставить жизнеспособность мух, гомо‑ и гетерозиготных по данной хромосоме. У гомозигот оба экземпляра второй хромосомы были идентичными, тогда как у гетерозигот они происходили из разных линий.
В начале опыта гомозиготы и гетерозиготы практически не различались по жизнеспособности, а затем во всех линиях начинался процесс постепенного «хирения» гомозигот, что можно объяснить накоплением второй хромосомой спонтанных мутаций с вредящим эффектом. Естественно, этот процесс шел во всех линиях совершенно независимо. Иногда случайно извлеченная хромосома счастливо избегала мутационных повреждений, в других случаях ей «везло» меньше. В некоторых случаях такая хромосома несла леталь, эти линии учитывались отдельно.
Усреднив полученные данные по всем линиям (свободным от леталей), можно было наблюдать, как падает жизнеспособность у гомозигот по хромосоме, фактически выведенной из‑под контроля очищающего отбора. К 60‑му поколению она снизилась примерно в 2 раза. Мукаи установил, что это вызвано накоплением полудоминантных мутаций, каждая из которых снижает жизнеспособность мух всего на 2–4 %. Скорость их появления составляет 0,4 мутации на геном гаметы за генерацию. Такие мутации возникают в геноме в 20–30 раз чаще рецессивных с сильным повреждающим эффектом (леталей и полулеталей).
Вернемся к искусственному отбору. В его ходе некоторый признак, обычно второстепенный для выживания (по крайней мере в лабораторных условиях) становится главным при разрешении вопроса жизни или смерти особи. Цель селекционера – сдвинуть в определенном направлении ценный, с его точки зрения, признак. Стремясь к этой цели, человек совершенно бессознательно способствует фиксации аллелей, благоприятствующих тому или иному уровню развития признака. Но локусы находятся в хромосомах, поэтому фиксация определенных аллелей – это фиксация довольно протяженных сегментов совершенно конкретных вариантов хромосом. Поддерживая в популяции высокий уровень благоприятных аллелей, селекционер фактически выводит из‑под контроля естественного отбора сцепленные с ними хромосомные сегменты. Поэтому в них могут беспрепятственно накапливаться мутации, снижающие жизнеспособность и плодовитость. В природной обстановке очищающий отбор удалял бы эти мутации, но находящиеся рядом ценные для человека аллели их сохраняют. Поэтому, несмотря на прогрессивный однонаправленный сдвиг признака, в популяции наблюдается снижение главных показателей приспособленности. Возникает стандартная ситуация – цели человека входят в противоречие с естественным порядком вещей. В этих условиях определенную «помощь» популяции может принести кроссоверная рекомбинация, способная отделить аллели, ценные с точки зрения человека, от аллелей, вредных с точки зрения естественного отбора.
Анализ генофонда популяций в ходе длительной лабораторной селекции показывает, что в нем действительно накапливаются мутации, снижающие главные компоненты приспособленности. В то же время, в процессе селекции появляются и благоприятные мутации, которые и позволяют в конечном счете преодолеть негативные эффекты отбора.
Доместикация
К настоящему времени накоплен довольно обширный материал (исторический и археологический), который позволяет проследить за изменением строения животных и растений при их доместикаций и в ходе дальнейшей культурной эволюции. Лучше всего этот материал освещает процесс доместикации в одном из ее главных очагов – регионе, история которого обычно излагается в разделе «Древний Восток». Этот географический район включает в себя обширную зону, в виде полумесяца охватившую восточную часть Средиземного моря, и две примыкающие к ней полосы. Одна – более широкая – протянулась от истоков Тигра и Евфрата вдоль западных склонов гор Загроса до Персидского залива, другая, узкая, вдоль Нила.
На территории Древнего Востока примерно 9 тысяч лет назад произошло величайшее в истории человечества событие – переход от присваивающего хозяйства к производящему. Именно здесь человек одомашнил несколько видов животных и растений, которые и по сей день являются основными источниками его пищи.
Не следует полагать, что человек сразу же после одомашнивания животных приступил к отбору на улучшение их хозяйственно ценных признаков. Все было гораздо прозаичнее – в течение 3–4 тысяч лет животных скорее всего съедали без всякой браковки. Такое положение вещей, вероятно, сохранялось примерно до момента возникновения первых цивилизаций в Междуречье и долине Нила. Обширные храмовые и царские хозяйства создали основу для возникновения отраслей экономики, ориентированных на производство мяса, молока, шерсти. С данного момента можно говорить о начале сознательной селекции, которая привела к созданию первых пород домашних животных. Переход к этой стадии сопровождался существенными изменениями морфологии, что запечатлено древними художниками Шумера и Египта. Можно проследить, как бараны теряют свою роскошную гриву и винтообразные рога, а их уши повисают. Любопытно, что примерно в это же время повисают уши и у коз, а у собак, быков и мелкого рогатого скота появляются пегости. Несколько позже громадные рога быков заменяются рожками шортгорнов. Все эти события датируются примерно вторым тысячелетием до нашей эры.
Странный параллелизм в появлении признаков, отметаемых в природной обстановке очищающим отбором, можно приписать негативному эффекту искусственного отбора. Выше мы отмечали, что этот эффект можно в какой‑то степени уменьшить синтезом новых, рекомбинантных хромосом, соединяющих аллели, полезные для человека, с аллелями, повышающими уровень главных компонент приспособленности. В таком случае искусственный отбор мог бы привести к повышению вероятности кроссоверного обмена и к более равномерному распределению этой вероятности вдоль хромосом. Недавно обнаружено, что уровень кроссоверной рекомбинации у домашних животных действительно гораздо выше, чем у диких.
История домашних животных демонстрирует не только мощь искусственного отбора, но не в меньшей мере и процесс деградации структур, функция которых в новых условиях оказалась избыточной. Домашние животные сами не заняты поиском пастбищ и защищены человеком от хищников. Следовательно, по сравнению с дикими предками их существование в меньшей степени зависит от скорости и глубины обработки внешних сигналов. Все это означает снижение давления очищающего отбора, следящего за работой органов чувств и центральной нервной системы. В итоге у всех домашних животных по сравнению с их дикими сородичами зарегистрировано уменьшение объема головного мозга на 15–30 %. У многих видов уменьшен диаметр глазниц. Как правило, абсолютные размеры животных также снижены. Степень деградации соматических структур у крупного рогатого скота и лошадей достигла максимума в Центральной Европе к концу средних веков. Современные высокопродуктивные породы получены на основе фактически карликовых форм, оставшихся от средневековья.
Исключение составляют скаковые лошади и некоторые породы собак, при выведении которых человек вел отбор на максимальную скорость бега. В этом случае цели человека не слишком расходились с обычными «целями» естественного отбора. В итоге деградация лошадей (прежде всего, уменьшение их размеров) связана обратной зависимостью с уровнем развития государства и его военных институтов. В частности, с падением Рима начался процесс резкого измельчания лошадей.
Попробуем просуммировать некоторые особенности, отличающие домашних млекопитающих от их диких предков. Они включают в себя изменение характера окраски шкуры (прежде всего многочисленные пегости); потерю или редукцию рогов; уменьшение числа хвостовых позвонков; уменьшение размеров тела и уменьшение величины головного мозга и органов, связанных с восприятием внешних сигналов; вислоухость (т. е. неспособность двигать ушами). Все эти изменения свидетельствуют о деградации функций, повышающих выживание в естественной (дикой) обстановке. Пожалуй, только функция размножения была усилена в ходе доместикации, что и не удивительно, так как, стремясь увеличить продуктивность животных, человек автоматически отбирал особей, приносящих наибольшее число потомков.
Итак, сходный (конвергентный) характер морфологических изменений при доместикации эволюционно неродственных видов скорее всего обусловлен снижением давления очищающего отбора, чему в немалой степени мог способствовать и интенсивный искусственный отбор, направленный на улучшение хозяйственно ценных признаков. Однако существует еще одно объяснение этой конвергенции. Советский генетик Д.К. Беляев предположил, что главная роль в процессе одомашнивания принадлежит отбору животных на ручной (неагрессивный) тип поведения. В ходе долговременного отбора на ручной тип поведения у серебристо‑черных лисиц Д.К. Беляев с сотрудниками обнаружили немало наследуемых морфологических отклонений, вполне типичных для процесса одомашнивания.
По мнению ученого, главная особенность отбора на поведение состоит в том, что он ведет к изменению гормонального статуса животных, а это, в свою очередь, может явиться причиной глубокой и направленной реорганизации онтогенеза. Постулируется, что изменение уровня гормонов может привести к наследуемому изменению активности ряда генов. Набор этих генов не вполне случайный, что и объясняет параллелизм в появлении аномальных форм у разных видов в процессе доместикации. Заметим, однако, что данная концепция оставляет без ответа многие вопросы. Получается, что сдвиг гормонального статуса взрослого организма может вызвать направленное изменение активности вполне определенных генов в клетках зародышевого пути, причем это измененное состояние генов способно сохраняться у потомков. Тем самым фактически предполагается превращение средовой (модификационной) изменчивости в наследственную.
Дата добавления: 2016-01-26; просмотров: 3620;