Медленные сдвиги среды
Итак, судьба разобщенных барьерами частей вида определяется соотношением двух процессов – экологической дифференциацией популяций и разрушением изолирующих барьеров. Находясь в несколько отличных экологических условиях, популяции под влиянием естественного отбора «стремятся» к ним лучше приспособиться, т. е. по ряду локусов зафиксировать разные аллели. Но этому стремлению противодействует генный поток, который, «просачиваясь» сквозь несовершенные барьеры, может вносить в каждую популяцию неблагоприятные (т. е. менее отвечающие местным условиям) аллели. Таким образом, генный поток выступает здесь как фактор, понижающий приспособленность каждой популяции, что позволяет говорить о миграционном генетическом грузе.
Есть только одно средство для закрепления успеха экологической дифференциации разделенных популяций – они должны за время своей изоляции стать разными видами, т. е. заменить географический барьер генетическим. Постоянное «стремление» вида расщепиться на несколько дочерних, по существу, представляет собой попытку вида освободиться от миграционного генетического груза. Кроме того, на периферии своего ареала вид может сталкиваться с качественно иными условиями среды, адаптация к которым означала бы прорыв вида в новую экологическую область.
Обрисованная здесь картина выглядит довольно оптимистично, но не следует забывать, что все эти адаптационные процессы предполагает фиксацию новых аллелей по десяткам локусов. Выше мы убедились в том, что на это требуются тысячи и даже десятки тысяч поколений. Но самое главное состоит в том, что естественный отбор проходит без селекционера, а где гарантии, что коэффициент отбора способен сохранять на протяжении нескольких тысяч поколений хотя бы свой знак? Иными словами, для адаптивной эволюции вектор естественного отбора должен сохранять направление и интенсивность в течение огромных отрезков времени. А учитывая противодействие со стороны генного потока, действие естественного отбора должно распространяться на значительную часть ареала вида, желательно на весь его ареал.
Естественный отбор – это метафора, его давление можно понимать как постепенное изменение интенсивности каких‑то жизненно важных факторов в пространстве и (или) во времени. Например, если ареал вида соседствует с экологической областью, где уровень его абсолютной приспособленности на порядок ниже, то необходимо, чтобы такой контакт сохранялся в течение нескольких десятков тысяч поколений. Или же по всей территории, занятой видом, должно идти однонаправленное изменение какого‑то жизненно важного параметра. Для адаптации к такой ситуации необходимо, чтобы скорость изменения параметра была очень небольшой. Во всяком случае, время перехода от оптимальных условий к очень плохим должно составлять тысячи или даже десятки тысяч лет.
О значении стабильности среды для экологической дифференциации, по‑видимому, свидетельствует удивительное явление повышения численности видов большинства групп растений и животных в направлении от полюсов к экватору. Действительно, климат в тропиках гораздо стабильнее, чем в умеренных широтах. Достаточно вспомнить о недавнем оледенении, практически не отразившемся на животном и растительном мире экваториальной зоны.
Кроме того, многие жизненно важные параметры среды испытывают медленные колебания с периодом, исчисляемым миллионами и даже десятками миллионов лет. Так, за последние 220 млн лет климат на Земле сменялся циклически с теплого на холодный не менее 16 раз, примерно по 15 млн лет на цикл (рис. 12). Уровень мирового океана также подвержен циклическим изменениям. Только за последние 65 млн лет его уровень 8 раз изменялся на 100–250 метров. Еще более медленны, но более грандиозны орогенические (горообразовательные) циклы. В них можно выделить длительный геосинклинальный период с продолжительностью до 500 млн лет; за ним в течение примерно 50 млн лет идет период бурного поднятия коры, после которого в течение нескольких десятков миллионов лет может длиться возвращение к геосинклинальному периоду. Во время периода горообразования горы «вздымаются» со скоростью около одного миллиметра в год. В геосинклинальный период эрозия стачивает поднятия быстрее, чем они появляются. Осадки могут накапливаться в мелководных эпиконтинентальных морях со скоростью до одного километра за 10 миллионов лет. Все эти геологические явления, среди которых следует упомянуть еще и дрейф континентов, изменяют, характер морских и воздушных течений, а тем самым и климат многих областей.
Итак, данные геологии свидетельствуют о том, что на Земле действительно происходили и происходят очень медленные и однонаправленные изменения среды, захватывающие огромные территории. Характерное время таких изменений выражается миллионами лет.
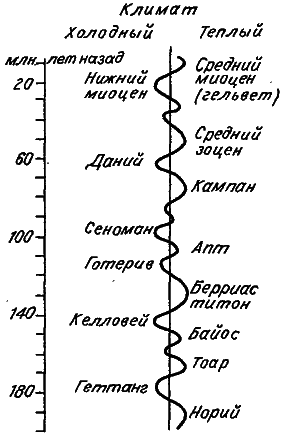
Рис. 12. Изменение среднегодовой температуры в течение юры и мела (по: [Красилов, 1977]).
Легко представить, что чем меньше ареал, занимаемый видом, тем больше у него шансов попасть под действие рассматриваемых здесь медленных однонаправленных сдвигов среды. Американский палеонтолог А. Буко проследил за изменением в течение силура и девона видового состава брахиопод, обитавших в разных геологических провинциях, по‑видимому, просто в разных эпиконтинентальных морях. Оказалось, что единственным фактором, от которого зависела скорость изменения фауны, была площадь этих морей. Чем меньше площадь, тем быстрее изменялся состав брахиопод. Заметим, что речь идет о миллиардах особей, занимавших ареалы в миллионы квадратных километров.
Рассмотрев в этой главе обстоятельства, при которых возможны ненейтральные изменения генофонда вида, мы приходим к двум важным выводам: во‑первых, уровень приспособленности вида к среде следует признать всего лишь удовлетворительным, и, во‑вторых, скорость фенотипического сдвига (приспособительного характера) не может быть очень высокой.
Дата добавления: 2016-01-26; просмотров: 803;