Отбор в панмиктической популяции
Перейдем к панмиктической популяции. Здесь при оплодотворении аллели гамет соединяются в генотипы зигот по правилам свободного попарного комбинирования. Зиготы с генотипами А1А1, А1А2 и возникают в соотношениях р2: 2pq: q2 (рис. 10). Эти же соотношения, получившие название закона Харди‑Вайнберга, будут справедливы и для взрослых членов популяции, если приспособленность всех генотипов одна и та же. Закон Харди‑Вайнберга довольно часто выполняется в природных популяциях, он позволяет легко вычислять частоты генотипов, исходя из частот аллелей.
В нашей модели отличия в приспособленности особей обусловлены только разным уровнем их жизнеспособности, поэтому численности зигот трех генотипических классов А1А1, А1А2 и А2А2 соответственно равны Np 2, 2Npq и Nq2. Тем самым зиготы в точности отражают генофонд популяции взрослых особей предыдущего поколения. В результате действия отбора численности генотипов взрослых особей, развившихся из зигот, приобретают вид
N11' = Np2w11; N12' = 2Npqw12; N22' = Nq2w22 . (3.10)
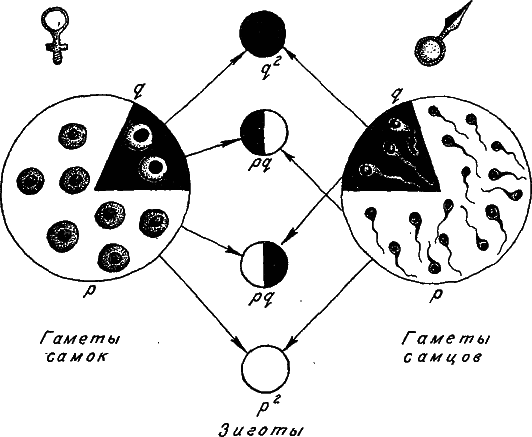
Рис. 10. Схема формирования генотипов зигот при случайном соединении гамет (при панмиксии).
Объяснения в тексте.
Численность всех взрослых особей после отбора связана с численностью зигот уравнением
N' = N(р2w11 + 2pqw12 + q2w22). (3.11)
Поскольку отношение N'/N выражает среднюю приспособленность популяции (см. уравнения (3.8)), поделив обе части уравнения (3.11) на N, получим
w = р2w11 + 2pqw12 + q2w22 . (3.12)
По аналогии с формулой (3.5) можно получить численность доноров аллеля А2 после отбора, т. е. среди взрослых потомков
N2' = N22' + 0,5N12' , (3.13)
или, учитывая соотношения (3.10),
N2' = Nq2w22 + Npqw12 . (3.14)
Поделив обе части этого уравнения на N2 = Nq (см. (3.6)), получим значение приспособленности доноров аллеля А2 :
w2 = N2' / N2 = (Nq(qw22 + pw12)) / Nq , (3.15)
т. е.
w2 = pw22 + qw12 .
Точно так же можно показать, что
w1 = pw11 + qw12 . (3.16)
Итак, мы располагаем всеми данными для вычисления сдвига частот аллелей после отбора (см. уравнение (3.9)).
Отбор в смысле Ч. Дарвина основан не на абсолютном уровне истребления молодых особей, а на различиях в степени такого истребления. В связи с этим приспособленность какой‑то группы особей удобно объявить равной единице, а приспособленность других групп представить в виде суммы 1 + s (s – коэффициент отбора). Такой прием позволяет рассмотреть наиболее интересную для нас задачу – судьбу мутаций, влияющих на относительную приспособленность их носителей.
Дата добавления: 2016-01-26; просмотров: 999;