Глава 3. Факторы микроэволюции
В природе существует много того, что не может быть ни достаточно глубоко понято, ни достаточно убедительно доказано, ни достаточно умело и надежно использовано на практике без помощи и вмешательства математики.
Ф.Бэкон
Для того чтобы в вопросах эволюции оставаться на твердой почве фактов, разумно руководствоваться принципом униформизма (актуализма), т. е. при объяснении явлений отдаленного прошлого исходить из анализа процессов, совершающихся в настоящее время. Не будем спешить и придумывать особые законы, влекущие эволюцию организмов по пути морфофизиологического прогресса. Сначала постараемся понять протекающую перед нашими глазами жизнь природных популяций, после чего сделаем попытку распространить эти (микроэволюционные) процессы на гораздо большие отрезки времени.
Генный поток
Сначала попробуем ответить на вопрос: что такое вид? Это понятие относится к наиболее важным и сложным в современной биологии. Начнем с того, что особи, связанные отношением родства, принадлежат одному виду. Под родством же люди обычно подразумевают каким‑то образом зарегистрированную общность происхождения. Например, братья и сестры имеют общими двух предков в поколении родителей. Двоюродные братья и сестры имеют общих предков в поколении дедов, троюродные – в поколении прадедов и т. д. Таким образом, с понижением степени родства общие предки перемещаются на все более отдаленные поколения. Совокупность особей, связанную отношением родства (включая и самые отдаленные его степени), можно было бы назвать видом. По существу, это очень близко к библейскому определению рода человеческого как всех потомков Адама и Евы. Однако такое определение крайне неудобно и ненадежно. Разве особи разных видов (например, волки и лисицы) не связаны отдаленным (эволюционным) родством? Как определить ту степень родства, которая еще позволяет отнести организмы к одному виду?
С генетической точки зрения, от предка к потомку передаются практически в неизменном виде наследственные факторы, гены. Поэтому общность происхождения – это общность генов. Миф об Адаме и Еве на самом деле довольно точно отражает реальную ситуацию, во всяком случае, есть веские основания считать, что вся совокупность наследственных факторов человечества восходит к генам очень небольшой группы особей. И если бы гены при передаче от родителей к потомкам не изменялись (т. е. не мутировали), то все люди были бы действительно братьями.
В ядре любой соматической (т. е. не половой) клетки организма имеются два аллельных варианта каждого гена, два аллеля – один от матери, другой от отца. Исключение составляют только половые клетки – гаметы, в их геноме каждый локус представлен всего одним аллелем, такой набор генов называется гаплоидным. При оплодотворении гаплоидные геномы гамет соединяются в диплоидный геном зиготы, который затем точно копируется механизмом клеточного деления. Таким образом, геномы всех соматических клеток организма на всех стадиях его развития (включая и зиготу) идентичны. Гаплоидный геном гамет возникает в результате действия особого клеточного механизма – мейоза. Суть его состоит в том, что в одну гамету попадает только один аллельный вариант каждого генного локуса, какой именно – материнский или отцовский – решает случай.
Сделаем одно важное (и в какой‑то степени спорное) предположение. Будем считать, что все особи одного вида обладают набором одних и тех же генных локусов, т. е. при мутировании может изменяться не число, а только аллельное состояние локусов. Потерю гена формально можно трактовать как его мутирование к нулевому аллелю. Правда, с добавлением гена (его дупликацией) вопрос обстоит сложнее. Итак, примем, что все особи одного вида содержат в хромосомах каждой клетки (в своем геноме) одно и то же число генных локусов, или генов.
Поскольку при передаче наследственного материала от родителя к потомку с некоторой вероятностью может произойти мутация по любому гену, то чем ниже степень родства особей, тем выше вероятность различия аллелей по каждому локусу. Различие аллелей ведет к различию фенотипов, т. е. к уменьшению внешнего сходства. Следовательно, степень родства можно оценить по близости внешнего вида особей. Мы оцениваем такое сходство на глаз, интуитивно схватывая огромное число признаков. В итоге, глядя на окружающие нас живые организмы, мы можем рассортировать их по внешнему виду, что, как правило, эквивалентно разбиению по видовой принадлежности.
Нет ли способа объективно оценивать степень сходства генов? Оказывается, такой способ есть, и им уже давно пользуются биологи. Считается, что особи разного пола принадлежат к одному виду, если они способны к самостоятельному скрещиванию и их потомство плодовито. При оплодотворении две гаметы родителей сливаются в зиготу, при этом объединяются два набора аллелей – от отца и матери. Под контролем конкретной комбинации аллелей (генотипа ) зигота превращается во взрослый организм. В связи с этим можно говорить о генетической программе развития организма, хотя это выражение не более, чем метафора, во всяком случае данная «программа» не имеет ничего общего с компьютерной. Итак, онтогенез потомка протекает под контролем сразу двух генетических программ. Если они не очень хорошо согласованы, то онтогенез может либо вовсе не завершиться, либо привести к созданию дефектной особи. Таким образом, метод скрещивания фактически оценивает близость аллелей по локусам, контролирующим онтогенез. Именно этим методом и пользуются особи одного вида в своей естественной жизни. Впрочем, для многих животных со сложным поведением оценка видовой принадлежности по внешним признакам также является необходимым условием для скрещивания, и спаривания не произойдет, если особи разных полов не «поймут» ритуала ухаживания.
Ситуация, когда любые две взрослые разнополые особи одного вида могут стать участниками скрещивания, называется панмиксией. Естественно, в реальности условия для панмиксии выполняются далеко не всегда, поскольку для соединения любой пары гамет нужно, Чтобы родители – обладатели гамет – находились недалеко друг от друга. Тем не менее в пределах этого «недалеко» панмиксия выполняется. Например, жителей большого этнически однородного города с хорошо организованным транспортом можно считать связанными панмиксией.
У большинства зоологических видов особи способны к активному движению, кроме того, могут двигаться их личинки или гаметы. У ботанических видов к широкому расселению способны семена, споры, пыльца, а нередко и фрагменты растений. Мы можем взглянуть на это явление немного иначе и увидеть в нем блуждание генов по ареалу вида. Данный процесс имеет много общего с диффузией броуновских частиц. Поток таких частиц в каком‑то направлении пропорционален градиенту (перепаду) их концентрации вдоль этого направления и коэффициенту диффузии – параметру, отражающему подвижность самих частиц. В конечном счете концентрация частиц во всей области их блуждания выравнивается, и наступает диффузионное равновесие.
Точно так же по ареалу вида «движутся» и аллели. Как и в случае диффузии, через некоторое время, определяемое размерами ареала вида и способностью генов к перемещению, должно установиться равновесие аллельных частот. Если скорость движения генов высока, то процесс выравнивания этих частот идет быстро. При панмиксии, когда в скрещивание может вступить любая пара разнополых особей, аллельное равновесие устанавливается практически моментально, т. е. за одно поколение. Нередко мы имеем дело только с переходом к такому равновесию, когда на территории, занятой видом, обнаруживаются градиенты по частотам разных аллелей. Эти градиенты – клины – можно изобразить графически в виде семейства изолиний, как это принято для топографических карт. Вдоль такой изолинии частота аллеля постоянна. Где изолинии гуще, градиент частоты аллеля круче.
По аналогии с диффузионным потоком можно ввести понятие о генном потоке как о переносе аллелей через единицу длины изолинии. Ясно, что такой поток будет тем мощнее, чем круче градиент и чем больше коэффициент диффузии генов. Наиболее мощные генные потоки образуются, когда в какую‑то область ареала с высокой концентрацией определенного аллеля проникает группа иммигрантов с другой частотой того же аллеля.
Представим себе, что область блуждания броуновских частиц разделена на несколько отсеков перегородками с разной проницаемостью. Понятно, что если все частицы поместить в один из таких отсеков, то характер последующей диффузии будет существенно зависеть от геометрии расположения перегородок и проницаемости каждой из них. Точно так же территория, где обитают особи одного вида, далека от однородности. Здесь могут быть и реки, и горы, и луга, и леса, – и все это создает препятствия (барьеры) для генных потоков.
Ясно, что если вид занимает сильно пересеченную местность, то время выравнивания концентраций аллелей будет много больше, чем в случае однородного ландшафта. Более того, в реальной ситуации существует немало районов, отделенных от остальных частей ареала практически непроницаемыми барьерами. Представим ареал как замкнутое пространство, разделенное на ячейки барьерами разной проницаемости. Внутри таких ячеек препятствий для генного потока практически нет, поэтому аллельные частоты в них выравниваются сравнительно быстро. Во всяком случае, скорость этого процесса внутри ячеек на несколько порядков выше, чем на их границе. Ясно, что даже в соседних ячейках частоты одних и тех же аллелей могут резко различаться. Вот эту‑то часть вида, населяющую одну ячейку, и назовем популяцией. Каждая популяция может быть охарактеризована своим генофондом, т. е. составом аллелей каждого локуса. Популяцию, занимающую ячейку с непроницаемыми стенками, назовем изолированной.
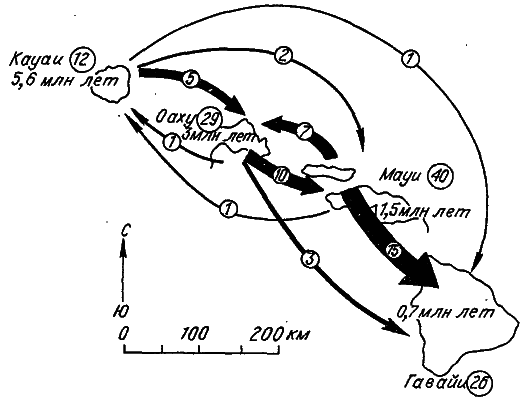
Рис. 9. Схема видообразования гавайских дрозофилид (по: [Carson, 1983]).
Стрелками показаны направление и число заносов мух‑основательниц новых видов. В кружках – количество видов‑эндемиков каждого острова.
Встает естественный вопрос, как особи попадают в изолированные ячейки? Можно лишь заметить, что абсолютно непроницаемых барьеров нет, кроме того, их проницаемость со временем изменяется. Если виды существуют в течение миллионов лет, то легко себе представить, что большинство барьеров в их ареалах изменяют за это время свою проницаемость в широчайшем диапазоне. Даже такой молодой вид, как Н. sapiens (его возраст оценивают всего в 40 тысяч лет), уже успел столкнуться с неоднократным наступлением и отступлением ледника, представляющего собой весьма эффективный изолирующий барьер.
Американский генетик Х. Карсон сумел восстановить родословную одной группы дрозофилид, обитающих на Гавайских островах (рис. 9). Эти вулканические острова возникали друг за другом в направлении с северо‑запада на юго‑восток. Самому древнему острову Кауаи 5,6 миллиона лет, а самому молодому острову Гавайи – всего 700 тысяч лет. Анализ политенных хромосом 97 видов гавайских дрозофил привел Карсона к заключению, что все они происходят от одного вида, обитавшего на острове Кауаи. Затем мухи проникли на более юный остров Оаху и, наконец, на остров Гавайи. Число родственных видов во много раз превосходит число островов, поэтому следует предположить, что иногда происходило попадание мух с одного заселенного острова на другой. При этом, несмотря на общность происхождения, иммигранты не скрещивались с туземцами, т. е. обе группы дрозофилид за время географического разобщения успевали стать разными видами. Залеты мух с острова на остров относятся к очень редким событиям, поэтому скорее всего в основе каждого нового вида стояла всего одна осемененная самка.
Этот пример показывает, что для интеграции генофондов популяций в единый генофонд вида продолжительность существования барьеров должна быть намного меньше времени превращения изолированной популяции в новый вид. Очевидно, что со временем в разделенных друг от друга частях некогда единой популяции могут произойти изменения аллельного состава генов, достаточные для того, чтобы воспрепятствовать спариванию особей или нарушить нормальное течение онтогенеза их потомков; иными словами, разобщенные генофонды станут несовместимыми. Такая несовместимость представляет собой особый, самый эффективный, генетический барьер, разделяющий популяции навсегда.
Итак, на популяции одного вида постоянно действуют два фактора, противоположных по своему конечному эффекту. Генный поток стремится слить генофонды популяций в единый генофонд, тогда как процесс видообразования пытается воздвигнуть между временно разобщенными популяциями непреодолимые генетические барьеры. Далее нам предстоит понять, какие факторы лежат в основе генетической дифференциации популяций.
Дата добавления: 2016-01-26; просмотров: 1171;