Соматосенсорная система в таламусе и коре
Среди сенсорных центров служащих анатомическим субстратом сознательного восприятия, главными являются те, которые расположены в таламусе и кope Они сгруппированы функционально в таламо-корти-кальную систему. [ Лемнисковая система имеет здесь особенно четкую соматотопическую организацию, так что мы можем говорить о проецировании или отображении периферии тела на таламус и кору. Неспецифическая система не обнаруживает такой точной соматотопической организации.
Таламус общий вид и подразделения. На рис. 2-13 представлен схематический, упрощенный вид таламуса правой половины мозга. Функционально его можно подразделить на следующие группы ядер: 1) специфические ядра сенсорных систем кожи, глаза и уха (красные); 2) неспецифические ядра (темно-серые); 3) моторные ядра (светлосерые); 4) ассоциативные ядра (белые).
Каждое из специфических ядер (красных) получает сигналы с периферии, которая хорошо определена анатомически и функционально. Там также выделяются прямые аксонные пути, идущие в кору и назад из коры. Специфические ядра зрительной и слуховой систем (соответственно латеральное и медиальное коленчатые тела, ЛКТ и МКТ, красные на рис. 2-13, А) рассматриваются в гл. 4 и 5. Специфическим ядром сома-тосенсорной системы является вентробазальное ядро. Его функциональные свойства обсуждаются ниже.
Неспецифические ядра таламуса, большинство которых занимает медиальное положение, включают (среди прочих) центральное латеральное и центральное медиальное ядра, обозначенные на рис. 2-13, А темно-серым; функционально,, они связаны с ретикулярной формацией ствола мозга (см. разд. 2.4).f Подобно ретикулярной формации, они по-
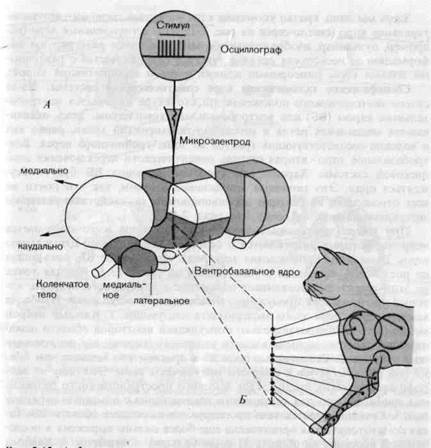
Рис. 2-13. А. Функциональные подразделения таламуса (схема). На схеме показан правый таламус; разрезы в двух местах представляют его внутреннее строение; показаны четыре группы ядер: специфические (красные), неспецифические (темно-серые), двигательные (светло-серые) и ассоциативные (белые). Микроэлек-трод, и дсцидлвг-раф--еимволизируют отведение потенциалов от одиночньпГнёй"-ронов вентробазального ядра. Ь. Участок длиной около 2 мм по ходу микроэлектрода через правое вентробазальное ядро кошки; показаны положения десяти нейронов, рецептивные поля которых обозначены на коже животного. (Poggio, Mountcastle, Bull. Johns Hopk. Hosp, 106, 266, 1960.)
лучают входные сигналы от всех сенсорных модальностей. Волокна, восходящие из спинного мозга, проходят преимущественно через передне-боковой пучок. Имеются эфферентные связи со многими областями коры, с гипоталамусом и лимбической системой. Неспецифические ядра таламуса являются, так._сказать, последующими станциями распределения афферентной информации, конвергирующей на ретикулярную формацию (рис. 2-7,Б и 2-12,Б)..
|
|
|
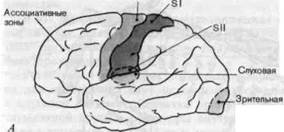
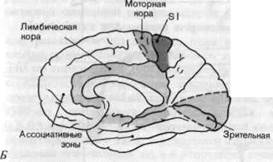
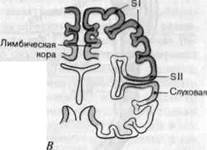
Рис. 2-14. Первичные сенсорные проекционные зоны коры. А. Внешний вид левого полушария. Б. Вид правого полушария со стороны срединной плоскости мозга. В. Фронтальное сечение левого полушария на уровне постцентральной извилины. Темно-красным показаны соматосенсорные проекционные зоны SI и SII, розовым-первичные проекционные зоны зрительной и слуховой систем. Кроме того, показано положение моторной коры (темно-серым), ассоциативных областей (белым) и кортикальных частей лимбической системы (светло-серым).
Здесь мы лишь кратко упомянем ядра моторной системы -вентролатеральное_ядро, (светло-серое на рис. 2-13, А) и ассоциативные ядра (например, пульвинар, изображенный белым); последние интегрируют информацию от нескольких органов чувств в сотрудничестве с различными зонами коры (сенсорными полями коры и ассоциативной корой).
Специфическое таламическое ядро соматосенсбрной системы. Из-за своего анатомического положения эта структура называется вентроба-зальным ядром (ВБ) или вентробазальным комплексом. Здесь оканчиваются медиальная петля и неоспинно-таламический тракт, равно как и волокна соответствующих трактов из зоны тройничного нерва. Вен-тробазальное ядро-вторая ступень синаптического переключения лем-нисковой системы. Характерные свойства нейронов ВБ будут обсуждаться ниже. Это типичные лемнисковые нейроны, так что почти во всех отношениях их реакции противоположны по свойствам реакциям экстралемнисковых нейронов (см. разд. 2.4).
При микроэлектродном отведении из ВБ чаще всего попадаются нейроны, которые разряжаются в ответ на механическую стимуляцию кожи. Траектория прохождения электрода через правое ВБ, показанная на рис. 2-13, А, приведена с увеличением на рис. 2-13, Б. Каждая точка н,а_этой траектории показывает положение в ядре одного нейрона, который реагировал на стимуляцию отмеченного участка кожи. Наиболее важные результаты этого эксперимента следующие. 1. Каждый нейрон может быть активирован только стимуляцией некоторой области левой передней лапы экспериментального животного (кошка); это рецептивное поле нейрона. 2. Рецептивные поля ВБ в среднем тем меньше, чем ближе они располагаются к опорным подушечкам лапы. Это одна из нейрофизиологических предпосылок высокого пространственного разрешения, необходимого для тактильного исследования с помощью передних лап. 3. Соседние участки тела проецируются в соседние области ВБ. Такая соматотопическая организация еще более сильно выражена в последующей проекции на область SI коры больших полушарий. 4. Каждый нейрон возбуждается только одним типом рецепторов-например, медленно адаптирующимися рецепторами давления или рецепторами кожных волосяных фолликулов (см. гл. 3). 5. Интенсивность периферического стимула кодируется средней частотой разряда нейрона, как это делается и в рецепторах (см. разд. 2.1).
Нейроны специфических таламических ядер глаза и уха (ЛКТ и МКТ) имеют аналогичные свойства.
Соматосенсорные проекционные зоны коры. Вентробазальное ядро (ВБ) таламуса связано восходящими и нисходящими аксонами с двумя областями коры, называемыми SI и SII (буква S ставится для обозначения сомашсёншрноыи, рис\ 2^14). SI располагается в постцентральной извилине, непосредственно за центральной бороздой-глубоким прогибом, проходящим через головной мозг. SII находится у человека на верхней стенке латеральной борозды, разделяющей теменную и височную доли.
Между периферией тела и контралатеральной областью SI имеются отчетливые соматотошческие взаимоотношения, проявления которых уже заметны и на более ниУКИХ уротпкпг=тт лемнисковых трактах и ядрах, и особенно в вентробазальном ядре таламуса (см. рис. 2-13, Б). На рис. 2-15 показано, как представлена на SI поверхность человеческого тела; это схематический рисунок, основанный на результатах электрической стимуляции коры у человека, находившегося в сознании. Геометрически искаженную проекцию поверхности тела на нейронную систему SI называют иногда «соматосенсорным гомункулусом». Эта j афферентная проекция напоминаеттшрту эфферентных связей моторной коры с мышцами.
Из рис. 2-15 ясно, что области коры, связанные с лицом и рукой, каждая в отдельности имеет примерно такую же величину, как проекционные области туловища и ноги взятые вместе. Это обще£_гхравил©4» органы с особенно высокой плотностью, рецепторов, такие, как пальцы,-й~гуоы7и проецируются соответственно на" самые "большие популяпии

Рис. 2-15. Соматотопическая организация зоны SI. Символически представленное отображение поверхности человеческого тела на постцентральную извилину, выявляющееся при локальном раздражении коры во время нейрохирургических операций (Penfield W., Rasmussen Т., 1950.)
нейронов SI. Следствия этого, касающиеся восприятия, будут обсуждаться ниже.
Как у животных, так и у человека соматотопическое отображение периферии тела на сенсорной коре можно изучать путем стимуляции периферии и регистрации в коре больших полушарий возникающих при этом вызванных потенциалов; латентные периоды между стимулами и вызванными потенциалами дают указание на число переключений в цепи. Таким способом были построены «карты» соматосенсорных
Проекционных ЗОН ДЛЯ МНОГИХ ВИДОВ ЖИВОТНЫХ. У высших приматов проекция SI похожа на-соответствующую проекцию у человека, показанную на рис. 2-15. У кошки и крысы непропорционально большое представительство имеют области рта и носа (в особенности тактильные вибриссы), тогда как передние конечности занимают относи-
тельно малую площадь по сравнению с тем, что характерно для приматов.
В SII также выделяется coмaтoтoпичecкaя организация, хотя и хуже выраженная, чем в SI особенностью S11 является то, что обе половины тела представлены в каждом полушарии, т.е. проекции являются
билатеральными.
Регистрация вызванных потенциалов может быть также использована для составления карт первичных проекционных зон в зрительной и слуховой системах. Сетчатка проецируется в ограниченную область затылочной доли, а рецепторы внутреннего уха имеют проекционные поля в височных долях. И те и другие показаны на рис. 2-14 (см. также рис. 4-32 и 5-16). Здесь также имеется поточечное представление периферической сенсорной поверхности на поверхность коры.
Переработка информации нейронами соматосенсорной коры.У нейронов, располагающихся прямо один над другим в 6 слоях S1. Рецептивные поля сильно перекрываются, тогда как у нейронов, лежащих рядом по горизонтали» они обычно четко различаются. Этот факт привел к заключению, что кора построена из функциональных единиц, представляющих-собой нейронные колонки, ориентированные перпендикулярно поверхности, причем каждая колонка имеет диаметр 0,7-0; 5 ммПри этом на нейроны каждой колонки могут влиять только рецепторы одного типа, т. е. колонки специализированы не только в отношении локализации, но и в отношении типа рецептогюв^Рднако в коре пока еще точно не установлено существование колонок, которые бы обрабатывали исключительно температурные стимулы, и специфических ноцицептивных нейронов в кортикальных проекционных полях пока еще тоже не обнаружено.
Теоретически каждая колонка коры благодаря большому числу содержащихся в ней нейронов (до 105) обладает значительными потенциями в смысле обработки информации с периферии. Было выдвинуто предположение, что колонки функционируют (на основе разнообразных возбуждающих и тормозных взаимодействий между нейронами) по иерархическому принципу; в этом процессе в результате соответствующих связей и операций разные свойства периферических стимулов кодируются разными нейронами. К этой функции ансамблей 'корковых "нейронов применяется термин выделение признаков. примеры, даны в разд. 4.3 для случая зрительной коры. После такой обработки информация направляется в другие области коры-например в моторную кору (прецёнтральная извилина) или в ассоциативные зоны.
Специфическая таламо-кортикальная система и сознательное восприятие. У человека и других приматов рука и область рта представлены в SI в особо крупном масштабе (рис. 2.15). Вместе с тем это именно те участки кожи, которые выделяются по своим тактильным способностям; пороги пространственного разрешения двух точек в этих областях особенно низки (см. рис. 3-3). Кортикальные проекции кожи, равно как и других органов чувств, принимают участие в процессе различительно-

 го, сознательного восприятия. В подтверждение этого свидетельствует целый ряд фактов, два из которых мы здесь опишем.
го, сознательного восприятия. В подтверждение этого свидетельствует целый ряд фактов, два из которых мы здесь опишем.
При хирургической операции на мозге, производимой под местной анестезией, больной способен сообщить о характере ощущений, возникающих при раздражении коры тонкими электродами. Например, если раздражению подвергается область кисти в зоне SI, больной ощущает прикосновение к кисти. Соматотопическая организация, представленная на рис. 2-15, была установлена именно таким способом. Значит, путем прямой стимуляции сенсорной коры можно вызвать сознательное ощущение, не активируя афферентные пути, включающие периферические нервы, спинной мозг и специфические ядра таламуса. Аналогичным образом стимуляция проекционных зон глаза производит ощущение вспышки света. Однако ощущение боли получить путем стимуляции коры не удается.
Экспериментальное удаление ограниченного участка SI у обезьян вызывает выпадение чувствительности в соответствующем периферическом участке. В одном опыте животное перед операцией научили отличать куб от шара на ощупь, не глядя на предмет. После удаления соответствующей руке сенсорной области коры эта способность оказалась утраченной ,и восстановить ее не удалось.
Аналогичные наблюдения были сделаны и на людях с повреждения-/ ми отдельных участков SI (например, в результате огнестрельного ранения или опухоли). Тактильные стимулы еще воспринимаются, но способность локализовать их нарушена; ощущения прикосновения имеют смутный, неопределенный характер. Боль чувствуется, но локализация болевых стимулов менее точна. Таким образом, интактность SI является предпосылкой пространственно организованного сознательного восприятия.
Электрическая стимуляция SIIу человека также вызывает ощущения, которые кажутся связанными с соответствующими участками периферии. Однако удаление только участков SII не вызывает отмеченных выше сенсорных выпадений. Роль SIIв сенсорном восприятии остается пока не выясненной.
Ассоциативные зоны коры.Все сенсорные проекционные зоны вместе с моторной корой занимают менее._ 20% поверхности коры (см. рис. 2-14). Оставшиеся области называются ассоциативными зонами. С_ первичными проекционными зонами их соединяют мощные тракты нервных волокон; обычно каждая ассоциативная область связана таким образом с несколькими проекционными зонами.
Термин «ассоциативная зона» происходит от функции, приписываемой этим участкам; полагают, что здесь осуществляются процессы ассоциации разнообразной, но специфической информации от органов чувств и формируются более сложные элементы сознания. В теменной ассоциативной области, расположенной между соматосенсорной и зрительной проекционными зонами, предположительно возникают наши субъективные представления об окружающем пространстве и нашем со-
бственном теле («образ тела»). Предполагается, что критическим моментом в формировании этих представлений является процесс научения в ходе развития в раннем детстве, во время которого оценивается и сопоставляется соматосенсорная, проприоцептивная и зрительная информация.
Гипотезы о функции ассоциативных полей у человека сформировались главным образом на основании наблюдения неврологических нарушений, возникающих в результате повреждения коры в ограниченных участках. Примером нарушения вследствие повреждения теменной ассоциативной области является зрительная агнозия.. У больного с таким синдромом зрение сохраняется в том смысле, что он может обходить препятствия или подходить к предметам и хватать их. Но он не может определять значение предметов. Когда больного, страдающего зрительной агнозией, просят назвать какую-нибудь вещь, на которую он смотрит, он не может этого сделать, но, взяв ее в руки, узнает ее с помощью осязания.
Относительная площадь, занимаемая ассоциативными зонами у приматов, и особенно у человека, намного больше, чем у других млекопитающих. Специфически человеческие ассоциативцый._ойдасциа расположенные в височной и лобной долях, Принимают участие в восприятии смысла речи и в формировании речи (речевые центры). Ассоциативным зонам посвящено очень мало нейрофизиологических исследований.
Экстралемнисковая система и сознательное восприятие.Возбуждение сенсорной коры-необходимое, но не достаточное условие появления сознательного восприятия. Многие клинические наблюдения и результаты экспериментов на животных свидетельствуют в пользу этого утверждения. Например, даже в условиях наркоза или глубокого сна можно вызвать первичные корковые потенциалы, но это не приведет к сознательному восприятию. Кроме того, повреждения верхней части ствола мозга, включающей ретикулярную формацию, приводят к стойкой потере сознания; электрическая стимуляция этой области у экспериментальных животных вызывает пробуждение, настораживание и переключение внимания. Два последних изменения поведения объединяют под названием «активация» (arousal). На основе этих и многих аналогичных данных возникло представление о том, что постоянный возбуждающий приток из ретикулярной формации в кору участвует в определении состояния сознания в диапазоне от глубокого сна через дремотное состояние до активного Водрстаования; появился термин «восходящая ретикулярная активирующая система».
Как схематически показано на рис. 2-16, сознательное восприятие, скажем, тактильного стимула, требует наличия двух входов в неокор-текс-специфической проекции по лёмнисковому пути (через вентробазальное ядро таламуса) И активациипосредством неспецифической системы ретикулярной формации (через неспецифические ядра таламуса). Лемнисковая система обеспечивает информацию об объекте восприятия (например, его положении, размере и времени действия), этот вклад


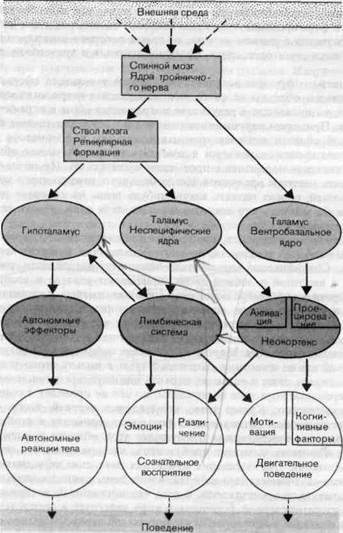
Рис. J--16. Взаимодействие специфической и неспецифической систем (очень упрощенная схема) в случае трех категорий поведения: сознательного восприятия, двигательного поведения и автономных реакций.
в сознательное восприятие обозначен на рис. 2-16 термином «различение». Вместе с тем неспецифическая активация, в которую могут внести вклад все сенсорные каналы, определяет уровень бодрствования как условие восприятия.
Но мы знаем из личного опыта (интроспекции), что каждое восприятие имеет еще один аспект-эмоциональный, или, как его обычно называют, чувственный: наше отношение к объекту восприятия. Эта размерность восприятия приписывается лимбической системе. Например, путем электрического раздражения области лимбической системы можно вызвать у кошки ярость и агрессивное поведение. Лимбическая система состоит из ряда подкорковых областей, грубо говоря располагающихся кольцом вокруг диэнцефалона.
Такая интерпретация ее функции предполагает, что лимбическая система вносит вклад в эмоциональное содержание восприятия. Как схематически показано на рис. 2-16, соматосенсорные входные сигналы поступают в лимбическую систему преимущественно по неспецифическому (экстралемнисковому) пути.
Лимбическая система имеет тесные реципрокные взаимоотношения с гипоталамусом - верховным управляющим центром для многих регу-ляторных цепей в автономной нервной системе (например, цепей регуляции сердечно-сосудистой активности и температуры тела) и эндокринной системе (например, цепи регуляции водного баланса). Возможность того, что сенсорные стимулы действуют на эффекторы автономных систем через гипоталамус, также учтены на схеме (рис. 2-16).
До сих пор, используя рис. 2-16, мы обсуждали только взаимодей- I ствия специфической и неспецифической систем. На том же рисунке i сильно упрощенно показаны взаимоотношения между тремя типами по-веденческих реакций на внешние стимулы. Это-сознательное восприя- ' тие, моторное поведение и бессознательные автоматические реакции тела.
Сознательное восприятие и двигательное поведение имеют по крайней мере две размерности, которые в каждом случае могут быть приписаны соответственно неокортексу и лимбической системе. Для случая восприятия мы уже обсуждали эти размерности - различение и эмоции. В двигательном поведении мы имеем, с одной стороны, когнитивные факторы (неокортикальные); они определяются конкретными сигналами от органов чувств. Вторая размерность в данном случае - это мотивация (drives); считается, что здесь участвует лимбическая система. Двигательные акты можно грубо подразделить на категории «приближение» и «избегание» в соответствии с тем, какая имеется тенденция-приближаться или удаляться.
Автономные реакции тела в значительной степени бессознательны. Их интенсивность, оцениваемая, например, по изменению кровяного давления или электрического сопротивления кожи, связана с интенсивностью эмоционального компонента восприятия. Сила мотивации дви- I гательного поведения также коррелирует с интенсивностью эмоций. «Общим знаменателем» здесь является возбуждение лимбической | системы. .
Графическое обобщение, представленное на рис. 2-16, следует рас- | сматривать как попытку изобразить то, что мы знаем о центральных
частях сенсорных систем, в виде наглядной схемы. Она обеспечивает читателю самую предварительную ориентацию в многочисленных сложных данных.
В 2. 16. Термин «соматосенсорная проекция» означает, что
а) тело нейрона посылает отростки в определенных направлениях;
б) периферическая сенсорная поверхность поточечно отображается на сенсорной коре;
в ) существует определенное представительство периферии на сенсорной коре;
г) нейроны соматосенсорной области коры SI связаны волокнами с нейронами ассоциативной коры. Верны несколько ответов.
В 2.17. Нейроны специфических ядер таламуса получают свои афферентные сигналы
а) от органов чувств, по крайней мере через семь синапсов;
б) от нескольких органов чувств одновременно, по крайней мере
через три синапса;
в) от терморецепторов и болевых афферентов кожи через ядра
задних столбов;
г) от одного органа чувств в каждом случае, хотя бы через два синапса. В 2.18. Какое из следующих утверждений приложимо к человеку, у которого разрушены области, соответствующие кисти и лицу в правой соматосенсорной коре SI?
а) Никакие объекты не могут быть распознаны на ощупь при исследовании правой рукой. V
б) Болевые раздражения левой руки ощущаются, но точность локализации снижена.
в) Отсутствует болевая чувствительность левой руки.
г) Благодаря билатеральной проекции в SII никаких заметных
нарушений не появляется.
В 2.19. Латеральное торможение в ЦНС ответственно за
v а) компенсацию пространственного распространения возбуждения, обусловленного дивергенцией; V б) уменьшение всех импульсов, вызываемых стимуляцией;
в) нейронный механизм усиления контраста;
г) подавление контралатеральных афферентных сенсорных
влияний.
Верны несколько ответов.
Дата добавления: 2015-10-05; просмотров: 3844;