Механорецепция
Этот раздел посвящен процессам обнаружения механических стимулов, действующих на кожу, и центральной обработке соответствующих сигналов, т.е. механорецепции (иногда называемой тактильной чувствительностью, или осязанием). Эта модальность объединяет четыре качества: ощущения давления, прикосновения, вибрации и щекотания. В дальнейшем, после обсуждения вопроса о субъективном восприятии механических стимулов, будет показано, что имеющиеся в коже различные механорецепторы обладают свойствами (специфика адекватных стимулов, адаптационное поведение), которые делают их особенно пригодными для восприятия того или иного из этих качеств.
Пороги ощущения и функция интенсивности в случае механической стимуляции кожи. Вэкспериментах, имеющих целью измерение порога для ощущения механического раздражения кожи, результаты сильно зависят от используемого метода стимуляции. При использовании самой старой процедуры, в которой пороги и распределение механочувстви-

Рис. 3-1. Пороги восприятия механического раздражения в различных участках кожи. А. Калибровка «волоска фон Фрея»; измеряется сила (в миллиньютонах, мН), при которой волосок или щетинка (нейлоновая) начинают изгибаться. Б. Пример механического стимулятора с электрически управляемым стержнем. В. Пороги ощущения в разных точках, выраженные через глубину прогиба кожи (в микрометрах, мкм); форма использованных стимулов показана на рис. 3-1, Б. (Lindblom, Lindstrom. In: Zotterman Y. (ed.), Sensory Functions of the Skin in Primates, Oxford, 1976.)
тельности определяются посредством волосков или щетинок, выясняется, что с помощью легкого надавливания (в диапазоне 0,1-0,5 г; метод калибровки представлен на рис. 3-1, А) можно вызвать ощущение давления или прикосновения (т.е. тактильные ощущения) только в определенных точках кожи. Эти точки называются тактильными точками. Результаты одного такого эксперимента показаны на рис. 3-18. Участки кожи с большим количеством тактильных точек-это кончики пальцев, и в особенности губы; на плечах, бедрах и спине тактильные точки особенно редки.
Эксперименты по определению тактильных точек кожи и порогового
давления в них информативны, но трудны и утомительны. Кроме того, у них есть тот недостаток, что из-за точечного характера стимуляции нелегко производить прямые сопоставления с обычно встречающимися стимулами, которые занимают ощутимую площадь. В современных стимуляторах, с помощью которых можно в широком диапазоне менять форму, длительность и интенсивность стимулов, используются сменные стержни, кончики которых контактируют с разными площадками на поверхности кожи. Посредством одного такого прибора при использовании формы стимула, показанной на рис. 3-1, Б, были получены значения порогов тактильной чувствительности на внутренней поверхности правой руки, представленные на рис. 3-1, В. Здесь пороги выражены через минимальную глубину прогибания кожи (в микронах), необходимую для появления едва заметного ощущения прикосновения.
Для появления тактильного ощущения на внутренней стороне руки достаточно вдавливания кожи на глубину порядка 0,01 мм (10 мкм). Пороги на кончиках пальцев отчетливо ниже, чем на остальной поверхности кисти. Этот результат согласуется с повседневным опытом. Однако тот факт, что пороги указательного и среднего пальцев не отличаются заметным образом от порогов других пальцев, кажется весьма удивительным. В том же самом исследовании-опять же довольно неожиданно-было обнаружено, что пороги тактильного ощущения у слепых не отличаются заметным образом от порогов у нормальных испытуемых.
Используя надпороговую механическую стимуляцию кожи, можно оценивать зависимость интенсивности ощущения от интенсивности стимула на основе психофизических методов, описанных в разд. 1.3. Пример данных, получаемых в таких исследованиях, приведен на рис. 3-2. В виде графиков, соотносящих величину действующего давления (абсцисса) с субъективно воспринимаемой интенсивностью стимула (ординаты), здесь представлены результаты для трех испытуемых (А, Б и В). Функции интенсивности были получены по три раза для каждого испытуемого; результаты ясно показывают, что эти функции хорошо воспроизводятся от раза к разу для каждого испытуемого, но сильно различаются у разных испытуемых. Этот факт отчетливо выражается в значениях показателей степенных функций, аппроксимирующих полученные данные. Следовательно, при оценке функций интенсивности такого рода (другие примеры см. на рис. 1-6 и 1-7) необходимо принимать во внимание не только условия эксперимента, но и индивидуальные различия в субъективных ощущениях.
Пространственные пороги различения.Используя пару ножек циркуля (с тупыми концами, чтобы избежать болевого раздражения), можно легко определять пространственные пороги различения, т.е. минимальные расстояния между двумя тактильными стимулами, при которых их удается воспринять как раздельные. Если оба кончика касаются кожи одновременно, т.е. исследуются пороги одновременного пространственного различения, на взрослых получаются результаты типа представленных на рис. 3-3. Они являются мерой пространственного разрешения

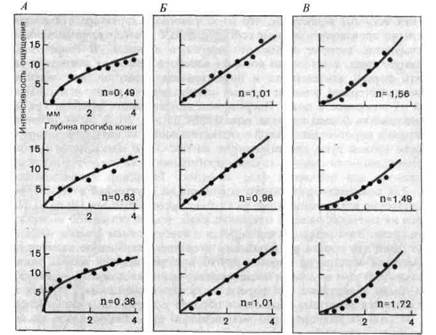
Рис. 3-2. Психофизически определяемые зависимости ощущения давления от интенсивности стимула. С помощью прибора, показанного на рис. 3-1, Б, производили раздражение ладони и измеряли глубину прогиба кожи; длительность каждого отдельного стимула составляла 1 с. Три испытуемых-Л, Б и В, давали численные оценки вызываемых этими стимулами ощущений. Каждый эксперимент повторяли 3 раза (соответственно в каждой колонке приведено по три графика). Сплошные линии-степенные функции, аппроксимирующие экспериментальные точки; показатели этих функций приведены на каждом графике справа. (Knibestol, Vallbo. In: Zotterman Y. (ed.), Sensory Functions of the Skin in Primates, Oxford, 1976.)
кожей тактильных стимулов в исследуемой части тела. Тот факт, что одновременные пространственные пороги кончика языка и пальцев, а также губ особенно низки (порядка 1-3 мм), тогда как на спине, плечах и бедрах они существенно выше (порядка 50-100 мм), согласуется и с повседневным опытом, и с описанным выше распределением тактильных точек. Отметим, кроме того, что вдоль линий, параллельных осям конечностей, эти пороги отчетливо выше, чем вдоль поперечных кольцевых линий (проверьте это на себе).
При прочих равных условиях пороги пространственного различения у слепых людей не ниже, чем у нормальных. Этот факт согласуется с отсутствием разницы в чувствительности к легким прикосновениям (см. выше); то же самое обнаружено и в случае вибрационных стимулов.
Если измерять пространственное разрешение в данной области ко-
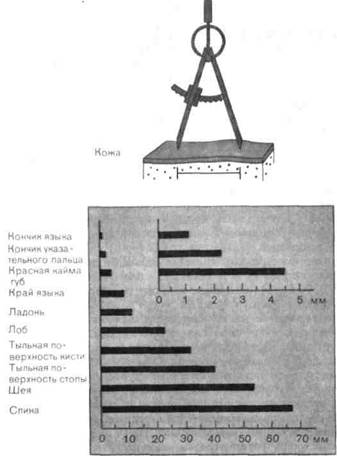
Рис. 3-3. Одновременные пространственные пороги у взрослых. На добавочном графике вверху справа в большем масштабе представлены три верхние полоски (увеличено в 10 раз). Длина полосок соответствует самому маленькому расстоянию между двумя стимулами, одновременно предъявляемыми в данном участке кожи, при котором эти два точечных стимула все еще воспринимаются как отдельные (Weber, Landois).
жи, прикладывая обе ножки циркуля последовательно, будут оцениваться не одновременные, а последовательные пространственные пороги. Если провести оба эксперимента на одном человеке, можно увидеть, что последовательные пространственные пороги явно ниже, чем одновременные, и часто составляют лишь четвертую часть от них (например, 1 мм вместо 4 мм). Причины этого различия отчасти кроются в механических свойствах кожи, но главным образом-в особенностях ее иннер-
вации и центральных связях афферентных нервных волокон (см. разд. 2.3).
Проверка механорецепции.При клиническом обследовании чувствительность к прикосновению обычно оценивают, прикладывая к коже клочок ваты или что-либо похожее и спрашивая пациента, что он чувствует и где ему кажется расположенным место стимуляции. Способность различать острое и тупое проверяют, воздействуя в случайной последовательности то острым кончиком, то головкой булавки со стеклянным шариком. Такие обследования также обычно включают выяснение у пациента того, может ли он узнавать цифры, выводимые на его коже каким-нибудь закругленным предметом, например пальцем или головкой булавки. При этом сначала изображаются крупные цифры, а затем-все меньше и меньше.
Чувствительность к вибрациипроверяется в клинике прикладыванием камертона к костным выступам (например, локтю или голени). В экспериментальных условиях или когда желательно более детальное обследование, лучше использовать электрический вибратор или громкоговоритель, приводимый в действие генератором синусоидальных колебаний. Целью, как правило, является нахождение абсолютного порога осознанного ощущения вибрации. Его минимум приходится на частоты около 150-300 Гц. Минимальные амплитуды вибрации, требующиеся на этих частотах, порядка 1 мкм.
Где только возможно, клинические измерения должны включать билатеральное сравнение -определение того же самого параметра на корреспондирующем месте другой стороны тела, причем следует обращать внимание даже на слабые различия. Обычное обследование во врачебном кабинете дает лишь относительно грубое представление о состоянии механорецепторной системы; слабые, но диагностически важные нарушения механочувствительности не всегда обнаруживаются.
Общие сведения о гистологической структуре, афферентной иннервации и адаптации кожных механорецепторов.На рис. 3-4 представлены виды механорецепторов, которые можно обнаружить у человека и других млекопитающих на не покрытых волосами (А) и на волосистых (Б) участках кожи. Все эти рецепторы иннервируются миелинизированными быстро проводящими афферентными волокнами группы II (диаметр 5-12 мкм, скорость проведения 30-70 м/с). При таких высоких скоростях проведения импульс, возникший на периферии, достигает спинного мозга за несколько миллисекунд.
В экспериментах на животных соотношение стимул-реакция для кожных рецепторов определяется путем регистрации потенциалов действия в одиночных афферентных нервных волокнах (рис. 3-5, А). Аналогичные отведения можно осуществить и на человеке, погружая через кожу в исследуемый нерв металлические электроды до тех пор, пока не будет уловлена активность одиночных нервных волокон. Путем стимуляции соответствующими приборами (см. рис. 3-1, А, Б) или прикладыванием грузов экспериментатор пытается вызвать раздражение рецепто-
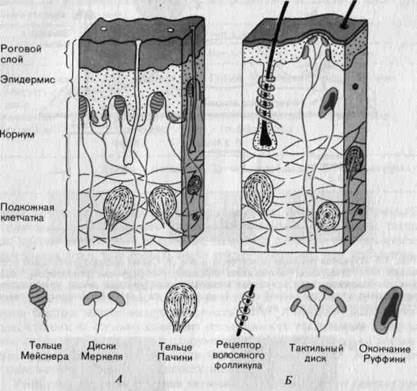
Рис. 3-4. Схема строения механорецепторов и их расположения в коже на не покрытых волосами (А) и волосистых (Б) участках кожи (см. также табл. 3-1).
ра, связанного с данным волокном, и затем определить зависимость между стимулом и реакцией.
При исследовании реакции на постоянное давление у различных рецепторов, показанных на рис. 3-4, становится очевидным, что по способности адаптации рецепторы могут быть разделены на три типа: адаптирующиеся очень быстро и реагирующие на каждый стимул только одним или двумя импульсами (тельца Пачини, см. рис. 3-8); адаптирующиеся быстро -разряд в них прекращается спустя 50-500 мс после включения стимула (рецепторы волосяных фолликулов, тельца Мейсне-ра; см. рис. 3-7), и такие, которые адаптируются медленно, продолжая генерировать вызываемые стимулом потенциалы даже тогда, когда давление поддерживается долгое время (тактильные диски, окончания Руф-фини, диски Меркеля; рис. 3-5, Б, Г и 3-6).
В табл. 3-1 представлена классификация рецепторов, основанная на этом критерии (адаптации к постоянному давлению). Данная классифи-
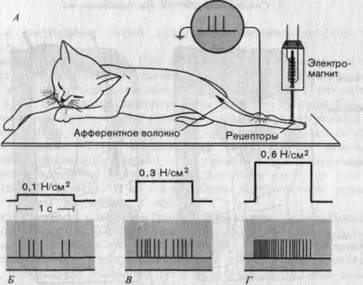
 Рис. 3-5. Изучение реакции рецепторов кожи. А. Схема экспериментальной установки. Б-Г. Импульсы (потенциалы действия), генерируемые рецептором давления, расположенным на подушечке кошачьей лапы (красные линии), при действии постоянных стимулов разной силы (в ньютонах на 1 см2). Спонтанной активности у данного рецептора не было.
Рис. 3-5. Изучение реакции рецепторов кожи. А. Схема экспериментальной установки. Б-Г. Импульсы (потенциалы действия), генерируемые рецептором давления, расположенным на подушечке кошачьей лапы (красные линии), при действии постоянных стимулов разной силы (в ньютонах на 1 см2). Спонтанной активности у данного рецептора не было.
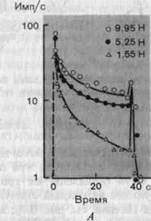
Рис. 3-6. Реакции рецептора давления на постоянные стимулы разной величины. А. Изменения разряда рецептора во времени (по ординате в логарифмической шкале отложено число импульсов в 1 с) при действии трех различных стимулов длительностью 40 с. Б. Зависимость между интенсивностью стимула и разрядом рецептора в разные моменты времени от начала действия стимула. По обеим осям масштаб логарифмический. Цифры на абсциссе соответствуют разностям между устанавливаемыми значениями интенсивности и пороговыми значениями для каждого момента времени. Каждая точка (на А и Б) представляет среднее из 10 отдельных измерений.
Таблица 3-1.Классификация кожных механорецепторов по скорости адаптации и адекватным стимулам
Адаптация к постоянному давлению
медленная быстрая очень быстрая
Участки кожи, не Диск Меркеля Тельце Мейснера Тельце Пачини
покрытые волосами
Волосистые участ- Тактильные диски, Рецептор волосяно- Тельце Пачини
ки кожи окончания Руф- го фолликула
фини
Датчик интенсив- Датчик скорости Датчик ускорения ности
кация возникла в то время, когда различные уровни интенсивности и длительности тактильного стимула устанавливать было легко, тогда как другие параметры стимуляции, например скорость или ускорение прогибания кожи, варьировать было трудно (например, при использовании волосков Фрея). Если рассматривать характеристики реакции по отношению к этим параметрам, оказывается (это будет показано ниже), что свойства рецепторов, адаптирующихся к давлению быстро или очень быстро, можно интерпретировать иначе. А именно: можно сказать, что они не столько кодируют интенсивность тактильного стимула (как это делают медленно адаптирующиеся рецепторы), сколько поставляют информацию о его скорости или ускорении. В табл. 3-1 это обстоятельство отражено в подписях под колонками.
Рецепторы давления (датчики интенсивности).У медленно адаптирующихся рецепторов мгновенная скорость разряда продолжает отражать интенсивность стимула на всем протяжении периода стимуляции давлением, даже если он очень длительный (рис. 3-5, Б, В, Г и 3-6, А). Если график зависимости между интенсивностью стимула и частотой разряда построить в логарифмическом масштабе (рис. 3-6, Б), то станет видно, что экспериментальные точки можно аппроксимировать прямыми линиями. Это означает, что данная зависимость есть степенная функция вида (частота импульсов) = (интенсивность стимула)" (см. разд. 2.1). С физиологической точки зрения такие рецепторы работают как датчики интенсивности, т. е. они измеряют силу или величину прогибания при действии механического стимула на кожу. Поскольку даже через длительное время эти рецепторы не адаптируются полностью (рис. 3-6), они также передают информацию и о продолжительности давления.
Медленно адаптирующиеся датчики интенсивности давления на участках кожи, не покрытых волосами (например, ладонная поверхность кисти), называют дисками Меркеля (рис. 3-4, А, табл. 3-1). Они располагаются небольшими группами в самых нижних слоях эпидермиса, откуда направляются отростки в сосочки собственно кожи-дермы. Диски
|
|
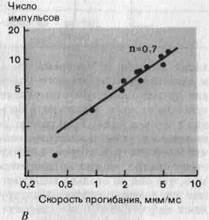
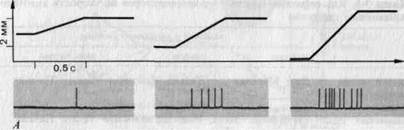
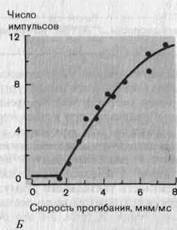
Рис. 3-7. Свойства реакций маханорецептора с быстрой адаптацией (датчик скорости). А. Реальные записи реакций (красные линии) на три механических стимула, различающихся скоростью прогибания кожи стержнем (отметки раздражения-черные линии), но действовавших в течение одного и того же времени. Б и В. Зависимость величины реакций рецептора от скорости прогибания кожи в линейном (Б) и логарифмичеком (В) масштабе. При пересчете в графике В из значений интенсивности вычитали величину порогового стимула (1,6 мкм/мс; Zimmermann).
Меркеля можно также обнаружить и на волосистых участках кожи, но там они располагаются в специальных тактильных дисках-небольших возвышениях кожи (рис. 3-4, Б). Кроме тактильных дисков в коже имеются также другие медленно адаптирующиеся рецепторы -окончания Руффини, располагающиеся в собственно коже (рис. 3-4, £>).
Рецепторы прикосновения (датчики скорости).Если на тыльной поверхности кисти отогнуть несколько волосков, не касаясь самой кожи, и оставить их в новом положении, что-то будет ощущаться только во время движения волосков. Как только они «замрут» в новом положении, это ощущение исчезнет. Данный факт указывает на то, что рецепторы волосяных фолликулов регистрируют не столько степень смеще-
ния волосков, сколько само их движение или, возможно, скорость движения.
В коже, не покрытой волосами, также имеются рецепторы, реагирующие сходным образом. Соответствующий пример приведен на рис. 3-7. Рецептор разряжается только во время непрерывного движения (наклонный участок на отметке) стимулирующего стержня; после его остановки разрядов нет, хотя стимуляция (прогиб) сохраняется. Частота импульсов явно зависит от скорости, с которой стержень погружается в кожу, что видно как по реальным записям (рис. 3-7, А), так и по графику (рис. 3-7, Б). Если эти данные представить в логарифмическом масштабе (В), то окажется, что график зависимости частоты разряда от скорости стимула (т.е. от первой производной по времени)-почти прямая линия; такая зависимость описывается степенной функцией. Следовательно, эти рецепторы можно назвать датчиками скорости (табл. 3-1).
Описанные выше датчики скорости-это тельца Мейснера, рецепторы с умеренной адаптацией, обнаруженные на не покрытых волосами участках кожи в сосочках дермы (рис. 3-4, А). На волосистых участках кожи их роль выполняют рецепторы волосяных фолликулов, располагающиеся на погруженных в кожу частях волосков (рис. 3-4, Б).
«Тонические» рецепторы, например рецепторы давления, называемые так из-за своего продолжительного разряда, сигнализируют преимущественно об интенсивности стимула. Соответственно «фазные» рецепторы ведут себя как датчики скорости; кроме того, имеются промежуточные (фазно-тонические) виды рецепторов. Иногда по аналогии с терминами, используемыми для технических датчиков, перечисленные рецепторы называют соответственно пропорциональными (П), дифференциальными (Д) и пропорционально-дифференциальными (ПД) датчиками.
Виброрецепторы (датчики ускорения).На рис. 3-8, А, Б показаны реакции рецепторов третьего типа при действии прямоугольных стимулов. Назависимо от силы стимула (стимул А - околопороговый, а стимул Б в несколько раз превышает пороговую интенсивность) рецептор генерирует только по одному импульсу на каждый стимул. Это значит, что он адаптируется очень быстро (табл. 3-1). Такой рецептор не может передавать информации ни о глубине прогиба кожи, ни о скорости прогибания.
Однако при синусоидальной стимуляции потенциалы действия вызывает каждая волна (рис. 3-8, В, Г). Если амплитуду синусоидальных колебаний установить на таком уровне, чтобы в каждый период возникало в точности по одному импульсу, станет ясно, что необходимая для этого минимальная амплитуда определенным образом зависит от частоты колебаний. При 110 Гц (Г) она меньше, чем при 55 Гц (В). На рис. 3-8, Д показана зависимость минимальной амплитуды колебаний, требующейся для возникновения на каждом периоде по одному импульсу (ордината), от частоты колебаний (абсцисса). Возрастание частоты стимуляции
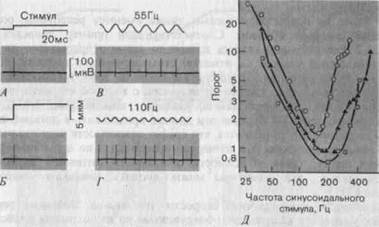
Рис. 3-8. Реакции телец Пачини (датчики ускорения). А-Г. Реакции на механические ступенчатые (А, Б) и синусоидальные (В, Г) стимулы; амплитуда синусоидальных стимулов находится вблизи порога генерации одного импульса на каждую волну; калибровка единая для всех записей. Д. Зависимость пороговой амплитуды от частоты при механических синусоидальных стимулах для трех телец Пачини. Стимулами воздействовали на места максимальной чувствительности каждого рецептора (подушечка кошачьей лапы). Обе шкалы-логарифмические.
с 30 до 200 Гц сопровождается резким снижением порога; на приведенном здесь графике в логарифмическом масштабе тангенс угла наклона кривой в диапазоне 30-200 Гц примерно равен — 2. Следовательно, зависимость пороговой амплитуды стимула S0 от частоты / можно записать так: S0 = const/"2, откуда можно заключить, что адекватным стимулом для этих рецепторов является вторая производная по времени от глубины прогибания, т. е. ускорение смещения кожи. В связи с этим мы можем назвать данные рецепторы датчиками ускорения (табл. 3-1). С увеличением частоты выше 200 Гц порог рецепторов этого типа возрастает, и при относительно высоких частотах (400-1000 Гц) реакцию типа 1 :1 получить уже не удается.
Датчики ускорения способны возбуждаться при стимуляции как волосистых, так и не покрытых волосами участков кожи. Как показано в табл. 3-1, на тех и других участках роль датчиков ускорения играют тельца Пачини, располагающиеся в жировой ткани подкожных слоев (рис. 3-4, А, Б). Это относительно крупные терминальные нервные образования, заключенные в слоистые оболочки из соединительной ткани, наподобие лука. Кроме подкожного жира они также обнаружены в разном числе на сухожилиях и фасциях мышц, на надкостнице и в суставных сумках.
Механочувствительные свободные окончания вкоже. Кроме миелини-зированных афферентов каждый кожный нерв содержит большое число
(50% и более) немиелинизированных аксонов (группа IV, или С). Некоторые из них-это эфферентные постганглионарные симпатические волокна, обслуживающие такие структуры, как гладкие мышцы сосудов кожи и волосяные фолликулы. Но в данную категорию входят также и афферентные волокна; эти нервные волокна имеют свободные окончания-на них нет корпускулярных структур. Рецепторные функции свободных нервных окончаний в ряде случаев пока неясны. Однако известно, что некоторые из них представляют собой терморецепторы, а многие -болевые рецепторы (см. разд. 3.3 и 3.4). Немногие из них чувствительны к слабым прикосновениям. Такие механорецепторы с немие-линизированными афферентными волокнами встречаются на волосистых участках кожи и изредка-на не покрытых волосами.
Из-за малой скорости проведения у волокон группы IV (порядка 1 м/с) между моментом стимуляции свободных нервных окончаний и прибытием афферентных импульсов в ЦНС проходит значительное время. Так, чтобы дойти по волокну группы IV от большого пальца ноги взрослого человека до спинного мозга (расстояние около 1 м), импульсу понадобится 1 с; тогда как импульсу, идущему по волокну группы II (скорость проведения 50 м/с), чтобы покрыть то же расстояние, потребуется всего 20 мс. Многие рефлексы, вызываемые механическими стимулами, и большая часть наших сознательных ощущений имеют более короткие латентные периоды, чем те, которые способны обеспечить волокна группы IV. Уже по одной этой причине данные волокна не могут играть роли в быстрых рефлексах, по крайней мере на их ранних стадиях.
Исследование особенностей реакций механочувствительных рецепторов, связанных с волокнами группы IV, обнаруживает еще одно важное их отличие от рассмотренных выше корпускулярных механорецепторов: кожные стимулы одинаковой интенсивности вызывают у них разные реакции. Это означает, что рецепторы данного типа не могут поставлять точных сведений об изменениях интенсивности стимула, т.е. в отношении оценки интенсивности стимула возможности одиночных механорецепторов, иннервируемых немиелинизированными нервными волокнами, очень малы. Они позволяют различать не более двух уровней интенсивности. Такая малая точность заставляет предполагать, что эти рецепторы являются пороговыми датчиками, которые сигнализируют только о наличии стимула в данном месте кожи. Кроме того, они могут принимать участие в передаче информации о слабых механических стимулах, движущихся по коже (например, о ползущем насекомом). Обсуждалось также предположение, что активация этих рецепторов, одних или в сочетании с другими, вызывает ощущение щекотки.
Рецептивные поля механорецепторов и плотность иннервации.Рецептивное поле механорецептора определяется как площадь, в пределах которой можно вызвать его возбуждение действием стимула определенной интенсивности; обычно используют интенсивность, в несколько раз превышающую интенсивность порогового стимула. Определяемое та-
ким образом рецептивное поле часто приблизительно соответствует анатомическим границам рецептора (это, например, справедливо для тактильных дисков). Однако в других случаях возбуждение рецептора могут вызывать и стимулы, действующие на некотором расстоянии (это, например, имеет место в случае телец Пачини). Сходным образом дело обстоит и с другими соматовисцеральными рецепторами.
Оценок плотности иннервации в различных участках кожи проведено немного, и те, как правило, косвенные (например, на основании измерений порогов различения). Кроме того, мы располагаем лишь разрозненными данными, касающимися дивергенции и конвергенции афферентных нервных волокон внутри рецепторных ансамблей. Например, известно, что на одно нервное волокно может приходиться по два-три тактильных диска, а внутри тактильных дисков все диски Меркеля (30-50 штук) обслуживаются одной нервной ветвью. У рецепторов волосяных фолликулов обнаружена более высокая степень дивергенции и конвергенции; тут одно афферентное волокно может обслуживать несколько сотен фолликулов, а каждый фолликул может иннервироваться множеством афферентных волокон. Общая картина усложняется значительными видовыми различиями в иннервации соответствующих участков кожи.
Данные об относительной встречаемости в коже различных механо-рецепторов также довольно скудны. Проведенные исследования указывают на то, что на волосистых и не покрытых волосами участках кожи обезьян и человека явное большинство составляют датчики скорости. С функциональной точки зрения это разумно, поскольку изменения ситуации для организма, как правило, важнее, чем абсолютные значения интенсивности и длительности стимулов. В этом смысле рецепторы скорости (фазные) иногда также называют детекторами новизны.
Свойства рецепторов и механорецепция.Разумно спросить, не является ли каждый из трех основных типов механорецепторов (подразумеваются датчики интенсивности, скорости и ускорения) ответственным за одно из трех различных ощущений-давления, прикосновения и вибрации соответственно. Тельца Пачини удовлетворяют такому предположению: при частотах стимуляции выше 60 Гц они, по-видимому, несут полную ответственность за ощущение вибрации. Но обычно встречающиеся механические стимулы содержат частоты ниже 60 Гц и поэтому раздражают одновременно несколько типов механорецепторов. На разные стимулы рецепторы этих типов реагируют по-разному, но результирующее ощущение нельзя связать с рецепторами одного типа. Соответственно в повседневной практике нам трудно определить различие между ощущениями давления и прикосновения.
Несколько лет назад, когда был разработан метод регистрации активности одиночных афферентных нервных волокон кожи человека с помощью металлических микроэлектродов, появилась возможность одновременно производить психофизические и нейрофизиологические измерения. Таким методом удалось показать, что очень слабые так-
тильные стимулы (с амплитудой порядка 4 мкм), которые при действии на кончик пальца возбуждают всего один рецептор с умеренной скоростью адаптации, вызывают некоторое ощущение. Однако на ладони пороги ощущения отчетливо выше, чем пороги самих механорецепторов (см. рис. 3-1, В). Это означает, что ЦНС «обращает больше внимания» на импульсы механорецепторов, если они поступают от кожных зон, более важных с точки зрения осязания.
В 3.1. Расположите следующие кожные зоны в порядке нарастания величины одновременного пространственного порога.
а) Край языка.
б) Красная кайма губ
в) Кончик указательного пальца.
г) Ладонь.
д) Спина.
е) Тыльная поверхность кисти.
В 3.2. Который из следующих терминов применим к тельцам Пачини?
а) Датчик интенсивности.
б) Датчик ускорения.
в) Пороговый датчик.
г) Датчик скорости.
В 3.3. Которое из следующих структурных образований имеет свойства датчика интенсивности?
а) Тельце Пачини.
б) Тельце Мейснера.
в) Диск Меркеля.
г) Рецептор волосяного фолликула.
В 3.4. Какого типа афферентные нервные волокна обслуживают рецепторы волосяных фолликулов?
а) Группа 1а.
б) Группа 16.
/ в) Группа II.
г) Группа III.
д) Группа IV.
В 3.5. Какие рецепторы из следующего списка являются датчиками скорости?
а) Тельца Пачини.
б) Тактильные диски.
b) Рецепторы волосяных фолликулов.
г) Тельца Мейснера.
д) Диски Меркеля.
В 3.6. Которое из следующих утверждений применимо к механочув-ствительным элементам с немиелинизированными афферентными волокнами (группы IV)?
а) Эти элементы имеют очень стабильную характеристику стимул-реакция.
б) Скорость проведения у этих афферентных волокон выше 2 м/с.
в) Число различимых градаций интенсивности больше трех.
г) Верны только утверждения бив.
д) Все утверждения (а-г) ложны.
Дата добавления: 2015-10-05; просмотров: 5308;