Проприоцепция
Всостоянии бодрствования мы отдаем себе отчет в ориентации наших конечностей по отношению друг к другу. Мы также воспринимаем движения наших суставов и способны довольно точно оценивать сопротивление любому движению, которое мы совершаем. Все эти способности вместе взятые называются проприоцепцией, поскольку источником эффективных стимулов во всех этих случаях служит само тело. Эта модальность имеет три качества - ощущения положения, движения и усилия; они будут рассмотрены ниже. Затем мы обратимся к соответствующим рецепторам-проприоцепторам, которые располагаются преимущественно в мышцах, сухожилиях и суставах. В конце будет показано, что структура осязаемого мира, равно как и нашего субъективного ощущения общего положения тела в пространстве,-это результат инте-гративной деятельности нервной системы, в которой важнейшую роль играют и другие сенсорные модальности, помимо проприоцепции, а также двигательная система.
Качества проприоцепции. Втемноте или при закрытых глазах мы представляем положение наших конечностей и ориентацию их частей относительно друг друга. Это качество проприоцепции называется ощущением положения. Строго говоря, это чувство информирует нас об углах в каждом суставе и тем самым-об относительном положении конечностей. Если мы не двигали конечностями долгое время или только что проснулись после долгого сна, наше ощущение положения конечностей обычно оказывается хорошо сохранившимся. Так что, по всей видимости, ощущение положения подвержено адаптации в слабой степени или совсем не подвержено.
Степень сгибания в суставе невозможно сколько-нибудь точно описать ни в градусах, ни в любой другой словесной форме без специальной тренировки. Однако с помощью следующих двух экспериментов мы можем легко продемонстрировать высокую точность, с которой мы воспринимаем относительное положение различных частей наших конечностей. Во-первых, любое положение одной конечности, в которое она была приведена экспериментатором или же произвольно самим испытуемым, последний может воспроизвести без зрительного контроля с помощью симметричной конечности. Во-вторых, мы можем, опять же без зрительного контроля, очень точно указать любую точку одной из конечностей (по собственному выбору или по желанию экспериментатора) при помощи пальцев кисти другой стороны.
Когда мы без зрительного контроля меняем суставной угол, например сгибаем или разгибаем руку в локте, мы воспринимаем и направле-
ние, и скорость движения. Это качество проприоцепции называется ощущением движения. Здесь порог восприятия зависит от величины и скорости изменения угла. Для проксимальных суставов (например, плечевых) этот порог заметно ниже, чем для дистальных (например, межфаланговых суставов пальцев).
Наша способность оценивать величину мышечного усилия, требующегося, чтобы совершить определенное движение или сохранить определенное положение сустава при действии какого-то сопротивления, также является одним из качеств проприоцепции. Мы называем его ощущением усилия. Поскольку сила, которую должна развивать мышца, зависит от препятствующего движению сопротивления, мы могли бы использовать в качестве синонима выражение «ощущение сопротивления», но этот термин не получил широкого распространения.
При экспериментальной оценке нашей способности определять усилие мы неизбежно наталкиваемся на трудности, связанные с исключением или учетом вклада кожных механорецепторов. Однако легко показать, что в случае ощущения усилия различительная способность явно лучше, чем в случае кожного ощущения давления. Вес какого-нибудь предмета намного легче оценить, если его приподнять, а не просто положить на кожу,-это факт, который знаком каждому из повседневной жизни.
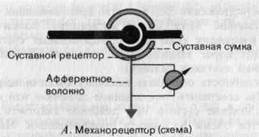
Проприоцепторы.Как отмечалось в начале этого раздела, первыми кандидатами на роль датчиков, обслуживающих данную модальность, являются рецепторы подкожных структур, называемые проприоцептора-ми. Главные из этих подкожных структур-мышцы, сухожилия и суставные сумки; все они содержат много механочувствительных элементов. В мышцах это в первую очередь мышечные веретена, а в сухожилиях -сухожильные органы Голъджи. Строение и реакции рецепторов этих двух типов подробно рассматривались в руководствах по нейрофизиологии; здесь они обсуждаться не будут. Однако в суставных сумках имеются многочисленные суставные рецепторы, которые мы сейчас опишем.
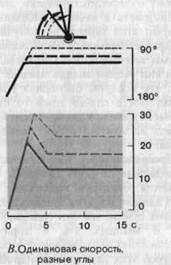
Поскольку при изменении положения сустава суставная сумка сжимается или растягивается, механорецепторы с подходящими свойствами могут сигнализировать как о положении сустава, так и о направлении и скорости движения, т.е. они могут определять ощущения положения и движения, но не усилия. Например, рецептор, схематически изображенный на рис. 3-9, А, расположен внутри суставной сумки таким образом, что сгибание растягивает его, а разгибание-сжимает. Графики реакций этого рецептора на сгибание сустава от 180 до 112,5° при трех различных угловых скоростях приведены на рис. 3-9, Б. Во всех трех случаях начальная фаза реакции рецептора состоит из серии учащающихся импульсов, причем частота их тем выше, чем больше скорость движения (сравните на 3-9, Б красные кривые для частоты разряда с черными, представляющими движение). После того как движение завершилось, рецептор начинает разряжаться с постоянной частотой, не

|
|
|
|
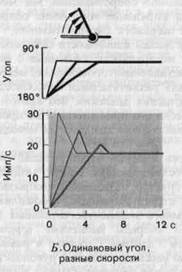
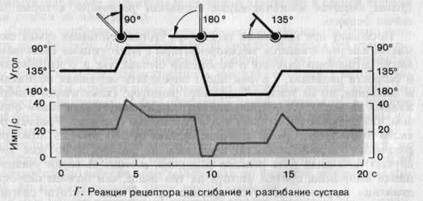
Рис. 3-9. Примеры реакций суставных рецепторов (упрощенные графики, основанные на данных Boyd, Roberts). А-Г. Объяснения в тексте.
зависящей от скорости предшествующего движения. Однако если данный сустав перевести с одной и той же скоростью в три различные конечные позиции, как это показано на рис. 3-9, В, то во всех трех случаях во время начальной фазы реакции частота разряда будет нарастать с одинаковой быстротой, а конечные частоты будут различными. Иными словами, частота разряда отражает как положение сустава по окончании движения, так и скорость движения (т. е. первую производную от координаты по времени). Другой примечательной особенностью, явствующей из рис. 3-9, Б, В, является то, что при определенном положении сустава разряд рецептора устанавливается на определенной частоте. Это означает, что после фиксации сустава в некотором положении рецептор еще долго посылает правильные сведения о том, какой угол образует сустав.
Фазно-тонические свойства суставных рецепторовпроявляются не только тогда, когда сустав движется так, что вызывает возрастание активности рецептора (как на рис. 3-9, Б, В), но и тогда, когда движение производится в обратном направлении. Это иллюстрирует рис. 3-9, Г на примере другого такого же рецептора. В исходном положении этот рецептор разряжался с частотой около 20 имп./с, что показано в левой части графика. Сгибание в суставе вызвало фазно-тоническую реакцию, подобную только что' описанной. Обратное движение-разгибание за пределы исходного положения (средняя часть графика)-вызвало отрицательную реакцию со столь сильным опусканием ниже начального уровня, что рецептор на время «замолчал». Возвращение к исходному углу (правая часть графика), сопровождавшееся второй положительной реакцией с «перехлестом» за начальный уровень, восстановило исходную частоту разряда.
Рецепторы, представленные на рис. 3-9, реагируют на сгибание увеличением частоты разряда, а на разгибание-уменьшением. Однако можно найти много других рецепторов, поведение которых будет зеркальным отражением этого. Сверх того, в очень подвижных суставах-таких, которые имеют много степеней свободы-можно также обнаружить рецепторы, избирательно чувствительные к вращению, отведению, приведению и т.д. Наконец, следует упомянуть, что в суставных сумках наряду с фазно-тоническими рецепторами имеются и фазные (быстро адаптирующиеся); последние разряжаются только во время движения.
У отдельных суставных рецепторов рабочий диапазон углов обычно меньше, чем это показано на рис. 3-9. Как правило, для перехода от нулевой активности к максимальной достаточно всего нескольких градусов, Достоинством такой особенности является то, что у отдельного рецептора на малые изменения угла приходится большая часть рабочей характеристики; это означает, что в пределах своего характеристического диапазона каждый отдельный рецептор будет регистрировать положение сустава с большой точностью. Такое подразделение всего диапазона движения сустава на перекрывающиеся зоны активности отдельных нейронов из некоторой популяции (называемое «дробление
 диапазона») дает гарантию того, что в ЦНС будет поступать точная информация о каждом положении сустава. Информация, посылаемая в мозг от многих рецепторов различных суставов, затем интегрируется, чтобы составилось общее представление об относительном положении суставов. Этот процесс интеграции, как и в случае других сенсорных сигналов, начинается в подкорковых сенсорных ядрах. Например, в таламу-се обнаружены нейроны, активность которых точно отражает положение сустава в диапазоне углов, превышающем 90°. Для этого там должна иметь место весьма точная и сложная конвергенция на такие нейроны волокон многих рецепторов из одного сустава, каждый из которых обслуживает малую часть общего диапазона движения.
диапазона») дает гарантию того, что в ЦНС будет поступать точная информация о каждом положении сустава. Информация, посылаемая в мозг от многих рецепторов различных суставов, затем интегрируется, чтобы составилось общее представление об относительном положении суставов. Этот процесс интеграции, как и в случае других сенсорных сигналов, начинается в подкорковых сенсорных ядрах. Например, в таламу-се обнаружены нейроны, активность которых точно отражает положение сустава в диапазоне углов, превышающем 90°. Для этого там должна иметь место весьма точная и сложная конвергенция на такие нейроны волокон многих рецепторов из одного сустава, каждый из которых обслуживает малую часть общего диапазона движения.
Гистологическая структура суставных рецепторов не вполне ясна. В суставной сумке имеются рецепторы типа окончаний Руффини (см. учебник гистологии), которые являются главными кандидатами на эту роль. Кроме того, там имеются рецепторы типа сухожильных органов Гольджи со сходным расположением внутри связок, и наконец, в меньшем числе там встречаются рецепторы типа телец Пачини. Несколько настоящих телец Пачини, как правило, обнаруживается в соединительной ткани, плотно облегающей сустав, но не в самой сумке. Все эти рецепторы обслуживаются миелинизированными афферентными нервными волокнами из групп II и III. Кроме того, там имеются свободные нервные окончания с немиелинизированными афферентами (волокна группы IV, или С), функция которых неизвестна. Похоже, что они принимают участие в передаче сигналов о болях в суставах.
Ощущения положения и движения могут также обеспечиваться кожными рецепторами в области суставов, так как при движениях суставов кожа сжимается или растягивается. Однако в опытах с применением местной анестезии таких участков кожи было показано, что кожные рецепторы играют в проприоцепции лишь второстепенную роль.
Центральная интеграция.По совокупности свойств суставных рецепторов можно сделать вывод о большой вероятности того, что в первую очередь именно они ответственны за ощущение положения и движения. Об этом свидетельствуют также и другие данные. Если вследствие патологии или местной анестезии передача по нервам суставной сумки прерывается, ощущения положения и движения сильно нарушаются.
Однако в восприятии положения и движения участвуют также и мышечные веретена; если у человека избирательно стимулировать эти рецепторы, воздействуя на сухожилия низкоамплитудной вибрацией (например, частотой 100 Гц и амплитудой в несколько микрометров), суждение испытуемого об истинном положении сустава искажается удивительным образом. Возникающие при этом ложные представления о положении можно выявить, если предложить испытуемому без зрительного контроля придать соответствующему суставу другой половины тела такое же положение, которое установил экспериментатор.
Наиболее вероятными датчиками для ощущения усилия являются рецепторы растяжения мышц-мышечные веретена и сухожильные op-
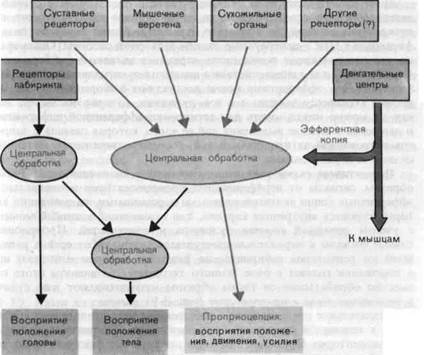
Vac. 3-10. Схема возникновения проприоцептивных ощущений. Чтобы обеспечить восприятие положения, движения и усилия, при обработке сигналов про-приоцепторов в нервной системе учитываются моторн"? 1ффрр'*цтчы(* у О""" В восприятие положения тела в пространстве наряду с проприоцептивной информацией вносит свой вклад информация, поступающая от рецепторов вестибулярного органа.
ганы. Следует, однако, помнить, что частота разряда афферентов мышечных веретен зависит не только от мгновенного значения длины мышцы, но также и от активности (а значит, и от напряжения) интрафу-зальных волокон, возбуждаемых моторными γаксонами. Было также показано, что у животных не удается вырабатывать условные рефлексы, используя избирательную стимуляцию волокон группы I. Исходя из этого следует предположить, что ощущение усилия определяется также и другими рецепторами, кроме уже упомянутых.
Таким образом, ни одна из рассмотренных рецепторных систем не способна независимо от других обеспечить информацию, необходимую для того или иного качества проприоцепции. Следовательно, в случае этой модальности восприятие требует одновременной активации различных рецепторных систем в специфических комбинациях и центральной интеграции таких афферентных сигналов (рис. 3-10).
Центральная интеграция включает также и использование сигналов несенсорного происхождения. Один из источников таких сигналов, составляющих важный, но еще мало изученный аспект переработки информации в ЦНС,-центральные отделы моторной системы. Оказывается, что они посылают копии своих «приказов мышцам» (эфферентные копии) в ЦНС для взаимодействия с сенсорными сигналами от проприо-цепторов. Эти эфферентные копии поставляют информацию о планируемой активности мышцы и о том движении, которое последует. Значит, их можно использовать для сортировки афферентной информации о движении в смысле выделения той ее части, которая связана с запрограммированной активностью, и той, которая определяется внешними силами и ограничениями.
Перцептивная схема тела и представление о положениитела. Таким образом, сигналы от периферических проприоцепторов и центральные эфферентные копии являются главными переменными, на основании которых строится внутренняя картина, или схема тела в данный момент с учетом движений головы, туловища и конечностей. Проприоцеп-тивные сигналы и параллельно поступающие сигналы от органа равновесия (от рецепторов лабиринта; см. разд. 6.1), которые извещают нас о положении головы в поле земного тяготения, в конечном итоге совместно обрабатываются таким образом, что позволяют нам судить о положении тела в пространстве (рис. 3-10, слева).
Сознательное представление о собственном теле удивительно крепко сидит в человеке и оказывается отделенным от афферентных сигналов проприоцепторов. Приведем в качестве примера тот факт, что после частичной и даже полной ампутации конечности подавляющее большинство пациентов продолжают ощущать отсутствующую конечность еще долгое время, часто до конца жизни. Эта иллюзия во многих случаях бывает столь убедительной, что пациент ощущает свою фантомную конечность более отчетливо, чем оставшуюся. Иногда пациент может по своему желанию двигать фантомной конечностью (влияние эфферентной копии ?); в других случаях он ощущает ее постоянно находящейся в неизменном положении. Часто случается, что с фантомной конечностью связываются соматосенсорные ощущения. К несчастью, многие из них неприятны, иногда-настолько болезненны, что синдром фантомных болей, которые не удается устранить никакими терапевтическими средствами, становится невыносимым для больного.
Структура осязаемого мира.Наше представление о пространстве формируется главным образом на основе зрительного восприятия. Но многие свойства нашего окружения лучше познаются, если провести исследование рукой - «потрогать и пощупать». Представьте, например, такие свойства, как жидкое, липкое, твердое, мягкое, гладкое, грубое, рифленое и многие другие, распознаваемые по комбинациям сигналов проприоцепторов, механорецепторов и кожных терморецепторов. Существенным моментом здесь является то, что при простом прикосновении (т. е. если мы просто положим предмет на руку или положим руку
на предмет и будем держать ее в одном положении) эти свойства плохо распознаются, если вообще распознаются. Однако, когда мы двигаем рукой, мы не испытываем затруднения в определении структуры и формы. Превосходство «ощупывающей» руки над неподвижной частично определяется активацией при движении значительно большего числа кожных рецепторов: адаптироваться рецепторы не успевают, и в ЦНС направляется более детальная информация о раздражителях кожи. Кроме того, при движениях руки в распознавание формы и упругих свойств ощупываемых предметов вносит свою долю проприоцепция.
В 3.7. Назовите три качества проприоцепции, о которых вы узнали в этом разделе.
В 3.8. Какой (ие) из следующих типов рецепторов в первую очередь ответствен (ны) за ощущение движения?
а) Рецепторы волосяных фолликулов.
б) Тельца Пачини.
в) Суставные рецепторы (типа рецепторов Руффини).
г) Рецепторы мышечных веретен.
д) Сухожильные органы Гольджи.
е) а, г и д совместно.
ж) в, г и д совместно.
В 3.9. Какие из следующих свойств тела легче определяются на ощупь, а не с помощью зрения?
а) Угловатый. г) Темный. ж) Жидкий.
б) Липкий. д) Мягкий. з) Яркий.
в) Круглый. е) Рифленый. и) Гибкий.
В 3.10. Какие из следующих типов рецепторов принимают участие в формировании представления о силе сопротивления? (Выберите три типа, которые вы считаете самыми важными.)
а) Диски Меркеля.
б) Рецепторы волосяных фолликулов.
в) Суставные рецепторы.
г) Тельца Пачини.
д) Рецепторы мышечных веретен.
е) Сухожильные органы Гольджи.
ж) Тактильные диски.
Дата добавления: 2015-10-05; просмотров: 3031;