Преобразование стимулов в рецепторах
Адекватный стимул, классификация рецепторов.Рецептор-это специализированная нервная клетка. Она сигнализирует в ЦНС о состоянии и (или) изменениях состояния среды, в которой находится. Те факторы окружающей среды и те их изменения, которые оказывают действие на рецепторы, называются стимулами. Стимулы-это величины, которые можно измерять объективными методами, например деформация кожи, температура и электромагнитное излучение (свет).
Физиологические исследования обнаружили, что каждый рецептор с особой готовностью реагирует на стимулы какого-то одного типа. Это свойство обычно называют специфичностью рецепторов; тот стимул, который является эффективным в каждом случае, иногда называют адекватным стимулом для данного рецептора. Для некоторых рецепторов адекватный стимул можно установить из повседневного опыта. Так, например, мы можем легко определить, что адекватным стимулом для рецепторов наших глаз является свет, тогда как тепловые или механические стимулы с помощью этих рецепторов мы в обычных условиях не обнаруживаем.
Следовательно, каждый рецептор способен передавать в ЦНС информацию только об одном определенном аспекте или размерности окружающей среды. Это значит, что рецептор играет роль фильтра. В связи с этим представляется полезным классифицировать рецепторы на основе их адекватных стимулов. Рецепторы млекопитающих распадаются на следующие" четыре группы: механо-, термо-, хемо-и фоторецепторы.
Внутри каждой из этих четырех групп мы можем усмотреть значительную степень специализации. Например, есть различающиеся типы фоторецепторов, по-разному отвечающие на излучения с различными длинами
волн; эти рецепторы называют чувствительными к длине во
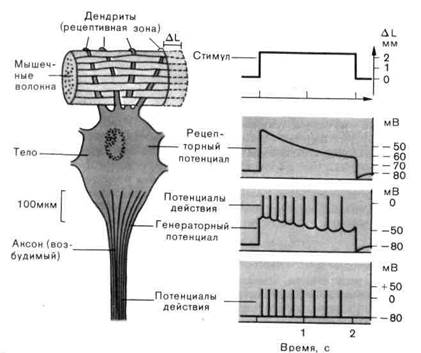
гас. г-1. схема электрофизиологических измерений, которые можно провести на рецепторе растяжения ракообразных. Во время стимуляции (изменение длины соответствующей мышцы на AL) микроэлектрод, введенный в тело клетки, регистрирует рецепторный потенциал, который возникает в рецептивной зоне (на дендритах). Являясь генераторным потенциалом, он возбуждает распространяющиеся потенциалы действия в возбудимом аксоне.
лны или цветочувствительными. Терморецепторы можно подразделить на рецепторы тепла и холода в соответствии с тем, что их возбуждает -увеличение или уменьшение температуры. У механорецепторов также можно наблюдать специализацию к различным параметрам стимулов: в коже, например, одни рецепторы чувствительны к вибрации, другие-к давлению.
Рецепторный потенциал. Вследующих параграфах мы рассмотрим процессы в рецепторе (подвергаемом действию адекватного стимула), результатом которых является потенциал действия в афферентном волокне. В качестве примера взят рецептор растяжения ракообразных-ме-ханорецептор, расположенный между определенными мышечными волокнами животного; он чувствителен к растяжению (рис. 2-1).
Когда микроэлектрод проникает в рецепторную клетку, то потенциал, который регистрируется исходно, как и во всех других нервных клетках, - это потенциал покоя (-80мВ на рис. 2-1). Под действием адекватной стимуляции - когда окружающая мышца растягивается мембранный потенциал меняется в направлении деполяризации. Эта вызываемая стимулом деполяризация-отклонение от потенциала покоя-называется рецепторным потенциалом. Рецепторный потенциал длится столько же, сколько и стимул (изменение ∆L на рис. 2-1). Однако даже тогда, когда стимул сохраняется постоянным, рецепторный потенциал снижается от своего исходного максимального значения до более низкого уровня. Такое снижение влияния стимула, поддерживаемого в течение некоторого времени на постоянном уровне, можно наблюдать практически у всех рецепторов; оно называется адаптацией.
Рецепторный потенциал в этом случае является результатом повышения проводимости мембраны, которое неспецифично и касается всех мелких ионов (Na+, K+, Са2+, Сl‾). В нормальных условиях единственный из этих ионов, который может обеспечить деполяризацию, это ион Na+, потому что только этот ион имеет потенциал равновесия, сдвинутый в деполяризационном направлении по отношению к потенциалу покоя. Следовательно, ионы Na+ должны быть главным источником возникновения рецепторного потенциала. Повышение проводимости мембраны, описанное для рецепторов, сходно с тем, что имеет место в субсинаптической мембране возбуждающих синапсов - например в концевой пластинке мотонейрона.
Повышение проводимости мембраны, которое вызывает рецепторный потенциал, локализовано в специализированном участке рецепторной клетки-рецептивной мембране (дендриты на рис. 2-1). Рецепторный потенциал возникает только тогда, когда стимул действует на этот специализированный участок. Далее, воспринимающий участок мембраны пространственно отделен от других частей клетки-например от электрически возбудимой (проводящей импульсы) мембраны аксона. Его отличие можно также выявить и фармакологически: например, возбудимую мебрану можно избирательно «отравить» нанесением тетро-дотоксина (ТТК), так что потенциалы действия больше там возникать не будут. В то же время рецептивная мембрана, а значит, и рецепторный потенциал, как правило, не чувствительны к действию ТТК.
Фоторецепторы позвоночных представляют исключение в том смысле, что при стимуляции светом в них возникает гиперполяризационный потенциал. Однако при синаптической передаче на нейроны высших уровней в сетчатке индуцируется деполяризация-возбуждающий пост-синаптический потенциал (ВПСП), который ведет к генерации распространяющихся потенциалов действия.
Процесс трансдукции; первичные и вторичные сенсорные клетки.Промежуточные события между поступлением стимула и появлением рецепторного потенциала пока еще мало изучены; они объединяются под общим названием процессы трансдукции. Этот термин подразумевает главным образом молекулярно-биологические процессы, имеющие место на рецептивной мембране и (или) во внутриклеточных структурах (называемых органеллами). В воспринимающей части рецептора имеется много таких органелл, как мембранные диски фоторецепторов, ко
|
торые содержат чувствительные к свету молекулы зрительного пигмента (см. рис. 4-27) или волосковидные выросты (реснички) рецепторов внутреннего уха (см. рис. 5-3). Это вспомогательные структуры, которые делают рецептор особенно чувствительным к специфическому адекватному стимулу.
Многие рецепторы состоят из двух связанных элементов: нервного окончания и некоторой клетки ненервного происхождения типа дисков Меркеля в коже (см. рис. 3-4) или чувствительных клеток во вкусовых почках (см. рис. 7-3). Полагают, что процесс преобразования сигнала имеет место в не-нервной клетке. Такие клетки называют вторичными сенсорными клетками. На электронной микрофотографии область контакта между клетками выглядит как синапс. По этой причине предполагается, что имеется синаптическая передача от вторичной сенсорной клетки к нервному окончанию.
Напротив, первичная сенсорная клетка -это рецептор, в котором преобразование стимула происходит в нервной клетке или в нервном окончании. Примером этого являются обонятельные рецепторы (см. рис. 8-2) и свободные нервные окончания в коже.
Рецепторный потенциал как причина генерации распространяющихся потенциалов действия.Рецепторный потенциал электротонически распространяется в смежные области клетки. В результате этого аксон деполяризуется (рис. 2-1). Когда деполяризация достигает порога мембраны аксона, генерируется потенциал действия. Этот потенциал действия проводится по аксону в ЦНС. Таким образом, рецепторный потенциал действует на аксон как электрический стимул; по этой причине его также называют генераторным потенциалом.
Если генераторный потенциал не спадает после генерации первого потенциала действия, он может вызывать следующие потенциалы действия, пока стимул и соответственно генераторный потенциал не исчезнут. Это значит, что в период существования длительного генераторного потенциала афферентное волокно разряжается многократно. Процессы, происходящие между поступлением стимула и генерацией распространяющихся потенциалов действия, могут быть названы процессами трансформации стимула в нервную активность. Таким образом, эта трансформация включает следующие события: трансдукцию, увеличение проводимости рецептивной мембраны, рецепторный потенциал, синаптическую передачу (в случае вторичных сенсорных клеток), генераторный потенциал, потенциалы действия.
Адаптация. При рассмотрении рецепторного потенциала мы отметили, что даже если стимул поддерживается некоторое время постоянным, рецепторный потенциал может уменьшаться (рис. 2-1); этот процесс называют адаптацией. Мы также наблюдаем этот феномен на картине разряда афферентных нервных волокон: как можно видеть из рис. 2-1, при неизменном стимуле временные интервалы ∆t между двумя последовательными импульсами удлиняются, т.е. мгновенная частота разряда уменьшается (мгновенная частота = 1/∆t). Адаптация может иметь место на всех стадиях трансформации стимула: в процессе трансдукции, в механизме проведения рецепторного потенциала, при синаптической передаче от вторичных сенсорных клеток и при генерации потенциала действия. Возможно, что несколько таких механизмов работает одновременно.

А Б
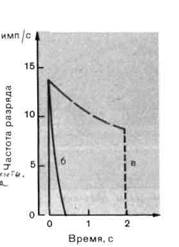
Рис. 2-2. Разряды рецепторов с различными скоростями адаптации. А. Потенциалы действия очень быстро (а), быстро (б) и медленно (в) адаптирующихся рецепторов, генерируемые при длительном действии постоянного стимула. Б. Графики изменения мгновенной частоты разряда со временем для рецепторов (б) и (в). Обратите внимание на то, что разряд рецептора (в) включает как фазиче-ский, так и тонический компоненты.
Чтобы получить адаптационную характеристику рецептора, исследуют его реакцию на надпороговый ступенчатый стимул. В таких экспериментах различные рецепторы демонстрируют различные изме
нения частоты разряда во времени. На рис. 2-2, А показан разряд медленно адаптирующегося рецептора (в), умеренно адаптирующегося рецептора (б) и быстро адаптирующегося рецептора (а). На рис. 2-2, Б представлено изменение со временем мгновенной частоты для рецепторов с медленной и умеренной адаптацией.
Предложены различные функциональные объяснения адаптации. Это явление обычно рассматривают всего лишь как простое утомление рецепторов. Но во многих случаях различное развитие адаптации во времени можно интерпретировать как приспособление к передаче специфических параметров стимула. Например, при изучении трех типов ме-ханорецепторов кожи было обнаружено, что благодаря характерным временным свойствам их разряда они могут рассматриваться как пере-

|
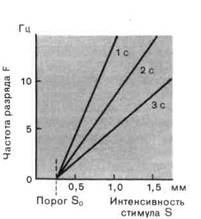 Рис. 2-3. Зависимость рецепторного потенциала и частоты разряда от интенсивности стимула. А. Рецепторные потенциалы и потенциалы действия при трех различных интенсивностях стимула. Потенциалы действия возбуждаются тогда, когда рецепторный потенциал превосходит порог (красная прерывистая линия). Б. Графики зависимости мгновенной частоты разряда F в разные моменты после включения стимула (через 1, 2 и 3 с) от интенсивности стимула S. S0 -пороговая интенсивность.
Рис. 2-3. Зависимость рецепторного потенциала и частоты разряда от интенсивности стимула. А. Рецепторные потенциалы и потенциалы действия при трех различных интенсивностях стимула. Потенциалы действия возбуждаются тогда, когда рецепторный потенциал превосходит порог (красная прерывистая линия). Б. Графики зависимости мгновенной частоты разряда F в разные моменты после включения стимула (через 1, 2 и 3 с) от интенсивности стимула S. S0 -пороговая интенсивность.
|
|
|
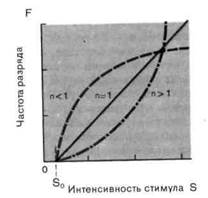
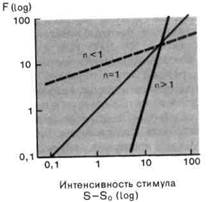
Рис. 2-4. Различные степенные функции. А. Частота разряда F как функция интенсивности стимула S. По обеим осям масштаб линейный, единицы-произвольные. Приведены три характерные степенные функции с показателями п < 1, п > 1 и п = 1 (красная линия). Б. Те же степенные функции изображены в логарифмическом масштабе. По абсциссе отложена разность между интенсивностью стимула S и пороговой интенсивностью S0.
| [2-1] |
датчики сведений об амплитуде, скорости и ускорении деформации кожи (см. рис. 3-5-3-8 и табл. 3-1).
Перевод интенсивности стимула в частоту импульсов в разряде.Когда сила стимула возрастает, возрастает и амплитуда рецепторного потенциала. Постепенные плавные изменения рецепторного потенциала вызывают соответствующие изменения частоты потенциалов действия. Эта ситуация представлена на рис. 2-3, А. Соотношение между интенсивностью стимула S и частотой импульсов F, часто называемое силовой функцией, изображено на рис. 2-3, £ для разных моментов времени от начала стимуляции (через 1, 2 и 3 с). В случае рецептора растяжения соотношение между F и S линейно для каждого момента времени на протяжении действия стимула.
Поскольку для того, чтобы вызвать потенциалы действия, рецепторный потенциал должен превзойти некоторое минимальное значение (порог аксона), разряд возникает только при превышении стимулом соответствующей интенсивности (S0 на рис. 2-3, Б). S0 называют пороговым стимулом для рецептора. Силовые функции рис. 2-3, Б описываются выражениями вида
F=k∙(S-S0),
т. е. линейной зависимостью для S > S0. Коэффициент к, наклон прямой, становится все меньше для последовательных интервалов времени, от-
считываемых от момента приложения стимула; это тоже является одним из выражений адаптации. '
Экспериментально было показано, что для большинства рецепторов силовая функция нелинейна. Измеренные соотношения часто можно описать различными математическими функциями, такими, как логарифмическая
| [2-2] |
F = k∙log S/S0
или степенная
F = k-(S-S0)ⁿ. [2-3]
Мы сталкивались с обоими этими выражениями в разделе по психофизике; они называются соответственно функциями Вебера-Фехнера иСтивенса. Кодирование интенсивности в рецепторах большинства типов лучше всего описывается степенной функцией (рис. 2-4), в которой разность- между интенсивностью стимула S и пороговой интенсивностью S0 возводится в n-ю степень. Показатель п является константой, характерной для каждого типа рецепторов. Для п < 1 и и > 1 степенные функции (на графиках с линейным масштабом) представляются соответственно выпуклыми и вогнутыми кривыми (рис. 2-4, А), а для п = 1 силовая функция линейна, как и для рецептора растяжения. Оказалось, что для большинства рецепторов и < 1. Силовая характеристика с п < 1
позволяет рецептору кодировать весьма значительный диапазон интен-сивностей стимула частотой разряда, ограниченной сверху (максимальная частота разряда толстого миелинизированного нервного волокна составляет 500-1000 имп./с, а у тонких волокон она еще меньше).
Степенные функции часто используются при описании психофизических результатов. Сравнение нейрофизиологически полученной силовой характеристики какого-нибудь рецептора с измерениями субъективного ощущения интенсивности в психофизическом эксперименте (см. рис. 1-6) часто обнаруживает поразительное соответствие; такие сопоставления важны для исследования нейрофизиологической основы восприятия.
Чтобы проверить, можно ли установленную в эксперименте силовую функцию описать степенной функцией, интенсивность стимула и частоту разряда представляют на графике в логарифмической системе координат (рис. 2-4, Б). В такой системе степенная зависимость выглядит как прямая линия, поскольку при логарифмировании обеих частей [2-3] получается
logF = logk + nlog(S-S0). [2-4]
Это уравнение прямой линии с угловым коэффициентом п для переменной logF при аргументе log (5 — S0). Если точки на таком графике могут быть аппроксимированы прямой линией, то они соответствуют степенной функции. Показатель п может быть определен непосредственно по наклону этой линии.
В 2.1. На какие четыре группы можно подразделить рецепторы млекопитающих по их адекватным стимулам? В 2.2. Рецепторный потенциал
а) это реакция рецепторной клетки типа «все или ничего», кото
рая не возникает до тех пор, пока стимул не превысит некото
рого порога;
б) обычно представляет собой деполяризацию рецептивной мем
браны, амплитуда которой нарастает с увеличением силы
стимула;
в) электротонически распространяется до мембраны аксона, где действует как возбудитель проводимых далее потенциалов действия;
г) возникает благодаря увеличению проводимости, связанной
с ионами Н+ (значением рН);
д) медленно нарастает при действии постоянного стимула
и длится в течение всего времени стимуляции.
Верны несколько ответов.
В 2.3. В афферентных аксонах многих рецепторов частота импульсов в разряде
а) увеличивается с увеличением интенсивности стимула;
б) увеличивается со временем при действии стимула постоянной интенсивности;
в) уменьшается со временем при действии стимула постоянной
интенсивности;
г) обычно равна нулю при действии стимулов подпороговой
интенсивности;
д) не зависит от величины рецепторного потенциала. Верны несколько ответов.
В 2.4. В случае многих рецепторов степенная функция описывает
а) временной ход адаптации;
б) временной ход рецепторного потенциала при увеличении силы
стимула;
в) зависимость частоты импульсации F от величины превышения
стимулом порога S0;
г) связь между амплитудой потенциала действия и величиной ре
цепторного потенциала.
Дата добавления: 2015-10-05; просмотров: 3572;