Свойства и функционирование сенсорных нейронов и нейронных ансамблей
На каждом уровне сенсорной системы поступление афферентных сигналов вызывает возбуждение и торможение отдельных нейронов, а также взаимодействие между нейронами и группами нейронов. Некоторые виды взаимодействий являются особенно характерными для ряда сенсорных систем. Они будут описаны в следующих разделах.
Латеральное торможение.На рис. 2-8, А показана нейронная сеть, состоящая из двух рецепторов и связанных с ними элементов на двух последующих синаптических уровнях. Обнаруженные здесь конвергентные и дивергентные нервные связи, как и везде в нервной системе, казалось бы, должны были приводить к лавинообразному распространению информации во все стороны на все более высоких уровнях ЦНС. Следовало бы ожидать, что представление точечного стимула в ЦНС будет с каждым синаптическим переключением все увеличиваться и становиться более размытым и менее точным. Однако с такими ситуациями мы встречаемся только в патологических условиях, например в случае отравления стрихнином. Стрихнин блокирует тормозные синапсы в ЦНС. Когда такое случается, малейший периферический стимул действительно порождает в нервных сетях цепные реакции, захватывающие обширные очаги. По-видимому, в норме торможение предотвращает такое катастрофическое распространение возбуждения.
Данный тормозный эффект базируется на характерной пространственной организации тормозных связей, нашедшей выражение в терминах латеральное торможение или торможение со стороны окружения; соответствующая иллюстрация дана на рис. 2-8, Б. На уровне первого синаптического переключения в афферентном пути каждый нейрон оказывает тормозное влияние на ближайшие к нему клетки посредством вставочных нейронов (на схеме они красные). Тот нейрон, который передает самый сильный афферентный сигнал (здесь это средний нейрон), оказывает и самое сильное тормозное влияние на соседей. Это ведет к снижению возбуждения в точках, удаленных от центра возбуждения. Таким образом, по сравнению с ситуацией, изображенной на рис. 2-8, А, пространственное растекание возбуждения будет снижено.
Было показано, что латеральное торможение существует на всех уровнях сенсорных систем; в соматосенсорной системе оно обнаружено в сером веществе спинного мозга, в ядрах дорсального столба, в тала-мусе и коре. Роль латерального торможения схематически представляют графики на рис. 2-8, Б. Латеральное торможение-свойство всех сенсорных систем, и особенно специфических. Его можно рассматривать как функциональную компенсацию растекания возбуждения, обусловленного нейроанатомией. Следствием этого является более четкая локализация информации о стимуле. Специфическое субъективное проявление латерального торможения-феномен усиления пространственного контраста в зрении (см. гл. 1).
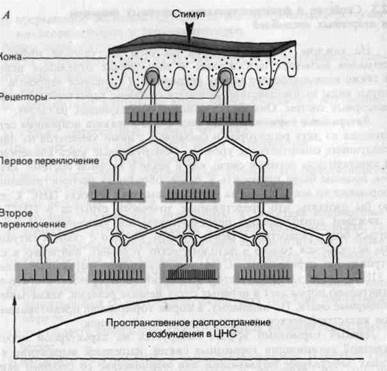
Рис. 2-8. Латеральное торможение. А. Пространственное распространение возбуждения в результате конвергенции и дивергенции в гипотетической ЦНС без латерального торможения. Вертикальные черточки (красные) символизируют импульсацию аксонов на различных уровнях. Кривая показывает соответствующее пространственное распространение возбуждения в ЦНС; эту кривую можно считать отражающей центральное представление стимула малой площади, показанного вверху.
Нисходящее торможение. Почти во всех сенсорных системах наблюдаются тормозные эффекты, порождаемые вышестоящими центрами. Такое нисходящее (центробежное) торможение может проявляться даже в таких периферических звеньях системы, как рецепторы (см. рис. 8-4) или афферентные окончания в спинном мозгу (рис. 2-10). Существование торможения такого рода подчеркивает важность сделанного выше замечания о том, что сенсорные системы не следует рассматривать как «улицы с односторонним движением».
Аналогично латеральному торможению нисходящее торможение можно считать средством регуляции чувствительности афферентных каналов передачи: вышестоящие части мозга «открывают клапан», управляющий потоком афферентной информации.
Рецептивное поле. Частота потенциалов действия сенсорных нейро-
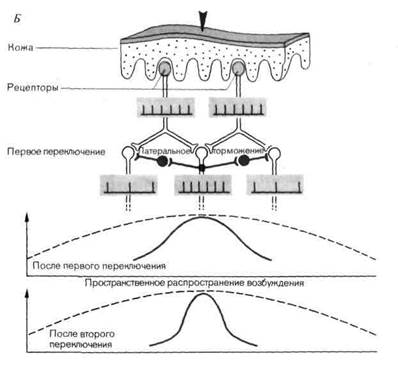
Б. Латеральное торможение (тормозное влияние каждого нейрона на соседние нейроны через тормозные нейроны; показано красным) на каждом синаптиче-ском уровне противодействует распространению возбуждения в результате дивергенции. Как показывают кривые распределения возбуждения, следствием этого является уменьшение области представления в ЦНС пространственно ограниченного периферического стимула.
нов обычно зависит от стимулов, действующих на периферии. Например, частота потенциалов действия какой-нибудь клетки зрительной коры может меняться, когда в определенной области поля зрения появляется определенный световой стимул. Совокупность всех точек на периферии тела, из области которых на данную клетку можно оказывать влияние действием специфических стимулов, называют рецептивным полем этой клетки. На рис. 2-9 приведен пример одного такого поля. На коже предплечья отмечены все точки, легкое прикосновение к которым может менять частоту разряда некоторой определенной клетки таламуса. Зона, внутрь которой попали все эти точки, обведена красным, она называется рецептивным полем таламической клетки для стимула «легкое прикосновение».
Некоторые клетки сенсорных центров имеют очень маленькие рецеп-


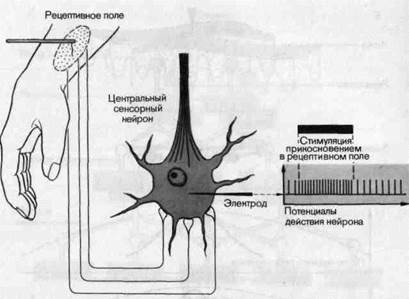
Рис. 2-9. Рецептивное поле центрального нейрона. Регистрируется активность центрального сенсорного нейрона; он генерирует потенциалы действия в ответ на прикосновение к коже руки в пределах области, показанной слева. Отметка раздражения и развитие реакции во времени показаны справа.
тивные поля. В качестве примера можно привести нейроны зрительной коры, которые реагируют на световые стимулы, действующие только в области желтого пятна глаза, имеющего площадь 0,02 мм2. Другие клетки, особенно принадлежащие к неспецифическим системам, представляют противоположный крайний случай: например, на такую клетку могут оказывать влияние стимулы, действующие на кожу обширной области тела, скажем всей ноги; причем эта клетка может реагировать и на прикосновение, и на охлаждение, и на вибрацию. В этом случае рецептивное поле имеет большую протяженность и охватывает различные модальности (полимодальная конвергенция).
Все, что отдельный сенсорный нейрон «знает» о положении специфического стимула,-это то, что стимул действует где-то внутри его рецептивного поля. Однако когда рецептивные поля нескольких нейронов одного уровня, которые во многих случаях значительно перекрываются, сравниваются на следующем, более высоком уровне, оказывается возможной более точная локализация стимула. В частности, такого рода переработка информации в нейронных сетях позволяет зрительной системе достичь высокой степени пространственного разрешения, несмотря на то что рецептивные поля отдельных нейронов в зрительных путях сравнительно велики (см. гл. 4).
До сих пор единственным критерием, по которому определялось ре-
цептивное поле, было «влияние» на данный центральный нейрон ст-Иму-ляции определенной точки на периферии. Но такие влияния могут быть как возбуждающими, так и тормозными, т.е. имеются возбуждающие и тормозные рецептивные поля или части полей. Для пример^ на рис. 4-28 показаны рецептивные поля сетчатки; в одном из случаев» возбуждающий центр окружен тормозной областью, в другом - ситуация обратная. Такое подразделение рецептивных полей может отражать нейронные связи, включающие латеральное торможение (см. в^,1ше)
Свойства рецептивного поля-
его положение и размер,модальности и качества специфических стимулов и подразделение на возбуждающуЮ и тормозную части-в принципе могут быть определены для каждой клетки сенсорной системы. Соседние клетки различных центров, как правило, имеют сходные рецептивные поля. Изучая синаптически связанные сенсорные нейроны на различных уровнях, можно выясните как происходит видоизменение рецептивных полей, и тем самым описать функциональные операции различных центров. От одного синаптическо-го переключения к другому размер и форма рецептивных полей могут меняться весьма характерным образом, а часто наблюдаются также и изменения вида стимулов, эффективных для разных уровней. Taк например , в зрительной системе для нейронов первого порядка в число существенных параметров стимулов входят такие , как яркость и цвет, а для зрительной коры стимулы могут быть более специализированными-скажем, нейрон может реагировать только на изменение Цвета.
Соотношения интенсивность-реакция, дифференциальные пороги,реакции центральных сенсорных нейронов можно охарактеризовать не только зависимостью от типа и локализации стимулов, но и зависимостью от интенсивности стимулов. Соотношение между частотой разряда нейрона F и величиной превышения стимулом пороговой интенсивности (S — S0) часто может быть описано степенной функцией
F = k(S-S0)
как это было обнаружено для многих рецепторов. У центральных нейронов также обнаружены различные значения п; встречаются и линейные соотношения (п = 1), но у многих нейронов наклон функции уменьшается при возрастании интенсивности стимула, т.е. показатель п меньше 1. Для центральных клеток также можно определять пороговые стимулы. Значение абсолютного порога S0 было включено в приведенную выше формулу степенной функции. Он измеряется путем нахождения наименьшей интенсивности стимула, на которую клетка реагирует изменением частоты потенциалов действия.
Порог другого типа можно определить, измеряя величину, на которую нужно изменить интенсивность надпорогового стимула, чтобы вызвать изменение частоты потенциалов действия нейрона. Такой порог называется дифференциальным порогом. Обнаружено, что он регулярным образом изменяется с изменением интенсивности стимула в со-
ответствии с правилом Вебера, описывающим дифференциальные пороги для субъективного ощущения.
Дифференциальные пороги можно определить и для многих других параметров стимула, помимо интенсивности,-пространственных и временных различий, различий в тоне и частоте выбрации. Для каждого случая приведенное выше определение справедливо; дифференциальный порог-это наименьшее изменение параметра стимула, которое вызывает измеримое изменение частоты разряда сенсорного нейрона.
В 2.10. Какое из следующих описаний рецептивного поля является правильным?
а) Правое полушарие мозга с полем зрения левого глаза.
б) Все точки на сетчатке, освещение которых ведет к увеличе
нию частоты импульсов в разряде некоторого нейрона зри-
^ тельной коры.
в) Все точки на сетчатке, освещение которых ведет к изменению
частоты импульсов в разряде некоторого нейрона зрительной
коры.
г) Область слизистой оболочки носа, которая может влиять на
активность некоторого нейрона обонятельной доли, если ис
пользовать локальное нанесение слабого раствора сероводо
рода.
д) Все точки на коже ноги, воздействие на которые болевым
стимулом может вызвать отдергивание ноги (сгибательный
рефлекс).
Верны несколько ответов.
В 2.11. Напишите по памяти определение дифференциальных порогов для нейронов сенсорной системы.
В 2.12. Предположим, что правило Вебера выполняется для стимула
«давление на кожу». При исходной нагрузке в 10 г дифферен
циальный порог некоторого нейрона равен 100 мг. Какова будет
величина дифференциального порога при исходной нагрузке
в 100 г? J
\ 1 2.4. Соматосенсорная система: спинной мозг, восходящие пути и ствол мозга
Связи афферентов в заднем роге спинного мозга. Вспинном мозгу аф-ференты из туловища и конечностей образуют синоптические связи с нейронами, лежащими в сером веществе.
Волокна от кожи и внутренностей заканчиваются на нейронах дор
сальных участков серого вещества-задних рогов (рис. 2-10). Часть
толстых (миелинизированных) афферентов (группа II) посылает допол
нительные веточки (называемые коллатералями), включающиеся в вос
ходящие пучки задних столбов. '
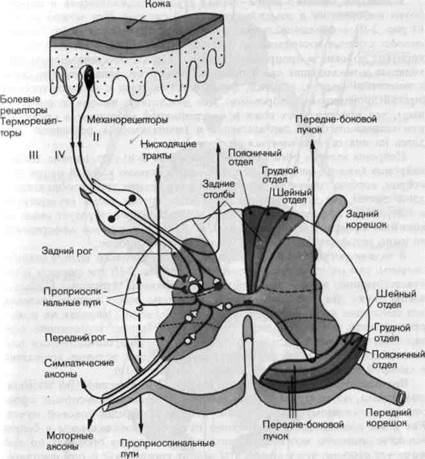
Рис. 2-10. Связи кожных афферентов в спинном мозгу. Афференты, входящие через задние корешки, образуют синапсы на нейронах заднего рога. Отсюда сигналы проводятся по волокнам, выходящим из спинного мозга (симпатическим и мышечным эфферентам), а также по восходящим путям. Из последних показаны пути, проходящие внутри передне-бокового пучка и ведущие в ствол мозга и таламус. Коллатерали афферентов группы II поднимаются в составе белого вещества (задних столбов) прямо в продолговатый мозг. Показаны два типа тормозных влияний на нейрон заднего рога (красные линии), они поступают по нисходящим путям и со стороны тормозных вставочных нейронов спинного мозга. Справа показаны слои, отражающие топографию восходящих путей на уровне шейного отдела.
|
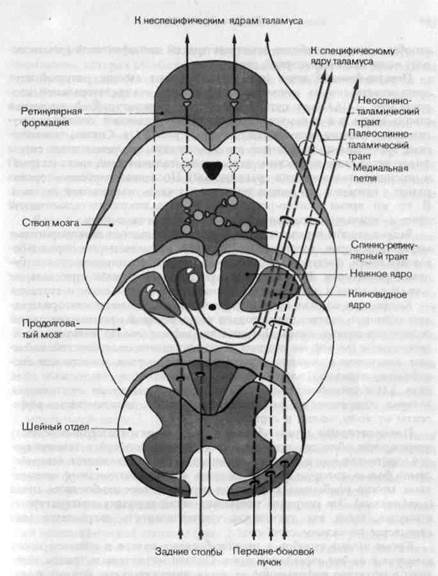
Рис. 2-11. Пути, восходящие из спинного мозга. Лемнисковые пути показаны красным, а экстралемниковые-черньш. Волокна задних столбов образуют синапсы в ядрах этих столбов (нежном и клиновидном) в продолговатом мозгу на нейронах, которые посылают свои аксоны в специфическое ядро таламуса через медиальную петлю. Филогенетически старые части передне-бокового пучка направляются в ретикулярную формацию (спинно-ретикулярный тракт) и неспецифические ядра таламуса (палеоспинно-таламический тракт). Филогенетически молодые части передне-бокового пучка (неоспинно-таламический тракт) проецируются в специфическое ядро таламуса; соответственно они рассматриваются как часть лемнисковой системы (красным
Задний рог спинного мозга-первая ступень переключения и переработки информации в соматовисцеральной системе. Как можно видеть из рис. 2-10, с функциональной точки зрения задний рог имеет четыре выхода: длинные восходящие пути (в частности, передне-боковой тракт), короткие волокна в проприоспинальных трактах, обеспечивающих восходящие и нисходящие связи соседних сегментов, синаптические связи с мотонейронами и, наконец, синаптические связи с симпатическими (преганглионарными) нейронами. Два последних выхода подразумевают, что афференты от кожи и внутренностей участвуют в организации спинномозговых двигательных и симпатических рефлексов. Мы здесь на них останавливаться не будем.
Нейроны заднего рога, показанные на рис. 2-10,-это многие тысячи, нейронов каждого сегмента спинного мозга, однако каждый отдельный нейрон, вообще говоря, не всегда имеет все четыре типа изображенных здесь связей. На этом рисунке дана лишь грубая схема конвергенции и дивергенции афферентных волокон. Каждый нейрон образует связи со многими афферентными волокнами, и, наоборот, каждое афферентное волокно синаптически возбуждает несколько нейронов.
В задних рогах спинного мозга имеются не только возбуждающие синапсы, там много также и тормозных. На рис. 2-10 эти синапсы и соответствующие вставочные нейроны изображены красным. Представлены также два типа торможения, имеющие особо важное значение для сенсорных систем,- афферентное и нисходящее. Основная их роль-регулировать и модулировать афферентный поток; торможение осуществляется отчасти постсинаптически, а отчасти пресинаптически (см. аксо-аксонный синапс на окончании одного из двух волокон, вошедших в спинной мозг через задний корешок на рис. 2-10).
Восходящие пути спинного мозга.На рис. 2-10 показаны два восходящих тракта, по которым информация идет от соматосенсорных аффе-рентов к головному мозгу,-задние столбы и передне-боковой пучок. Как и все восходящие и нисходящие пути, они локализованы в белом веществе спинного мозга. В задних столбах волокна проходят по той же самой стороне, что и афференты задних корешков (т. е. они ипсилате-ральны), тогда как в передне-боковом пучке-по другой стороне (кон-тралатерально).
Задние столбы состоят из прямых коллатералей толстых миелинизи-рованных афферентов задних корешков (групп I и II); все они без исключения приходят от низкопороговых механорецепторов туловища и конечностей-кожи, внутренностей, мышц и суставов. Все эти афференты, кроме мышечных, заканчиваются синапсами на нейронах ядер задних столбов (нежное и клиновидное ядра) в продолговатом мозгу (рис. 2-11).
Аксоны этих нейронов переходят на другую сторону и по ней идут в таламус в составе медиального лемниска (медиальной петли). Переработка нервных сигналов в ядрах задних стобов, как в заднем роге и та-ламусе, включает и афферентное и нисходящее торможение. Задние
 столбы и петля-наиболее заметные тракты специфической (лемниско-вой) части соматосенсорной системы.
столбы и петля-наиболее заметные тракты специфической (лемниско-вой) части соматосенсорной системы.
Передне-боковой пучок(рис. 2-11) содержит аксоны, которые идут преимущественно от нейронов заднего рога контралатеральной стороны (рис. 2-10). Этот путь заканчивается в ретикулярной формации ствола мозга и в таламусе; соответственно различают спинно-ретику-лярный и спинно-таламический тракты (рис. 2-11). Спинно-таламиче-ский тракт сам по себе имеет два компонента, различающиеся своим филогенетическим возрастом: палеоспинно-таламический тракт (старый) и неоспинно-таламический тракт (новый). Последний особенно хорошо развит у приматов; он рассматривается как часть лемнисковой системы. В то же время спинно-ретикулярный и палеоспинно-таламический тракты - важные экстралемнисковые спинномозговые пути.
Задние столбы передают сигналы исключительно от низкопороговых механорецепторов, тогда как информация, проходящая по передне-боковому пучку, поступает от механорецепторов, терморецепторов и болевых рецепторов. В оба тракта афферентные волокна приходят не только от кожи и внутренних органов, но также от мышц и суставов.
Когда один из этих путей перерезается, наблюдаемая сенсорная недостаточность соответствует только что описанной организации связей рецепторов разных типов. Повреждение заднего столба вызывает ипси-латеральное (на той же стороне тела) нарушение способностей, имеющих отношение к локализации тактильных стимулов, в частности способности отличать два точечных прикосновения от одного (см. разд. 3.1) и способности совершать исследование путем ощупывания, которая обеспечивается цепями обратной связи, включающими аффе-ренты от кожи, мышц и суставов.
После перерезки передне-бокового пучка в контралатеральных месту повреждения областях тела отмечается потеря болевой и температурной чувствительности. При заболеваниях, сопровождающихся непереносимой болью, которую не удается снять фармакологическими препаратами, можно прибегнуть к операции перерезки передне-бокового пучка (хордотомия). Эта операция прерывает также передачу температурных стимулов, тогда как тактильная чувствительность сохраняется (она передается по задним столбам).
Кроме задних столбов и передне-боковых пучков в спинном мозгу имеются и другие восходящие пути. Спинно-мозжечковые тракты передают в мозжечок информацию от механочувствительных кожных и мышечных афферентов; функционально они принадлежат к моторной системе. Спинно-шейный тракт, ведущий в таламус, несет преимущественно кожные афференты от конечностей. Его функция пока не ясна. Кроме длинных трактов, упоминавшихся до сих пор, существуют восходящие к головному мозгу полисинаптические пути, состоящие из последовательно соединенных нейронов спинного мозга с короткими аксонами.
Аксоны, выходящие из того или иного сегмента, идут внутри восхо-
дящих трактов бок о бок. Вследствие этого тракты имеют слоистую организацию, которая схематически представлена на рис. 2-10 для задних столбов и передне-бокового пучка на уровне шейного отдела. Когда аксоны входят в тракт, они поворачивают, чтобы идти параллельно волокнам из нижних отделов в части тракта, находящейся на стороне серого вещества. Границы отдельных пучков не так резко выражены, как это показано на рис. 2-10; в действительности имеют место диффузные переходы и перекрытия зон. В связи с такой топографической организацией при поверхностном повреждении передне-бокового пучка в шейной области (которое может быть вызвано травмой или опухолью) в первую очередь следует ожидать симптомов выпадения в нижней половине тела.
Ядра и восходящие тракты тройничного нерва.Тройничный нерв-со-мато-висцеральный сенсорный нерв лицевой области - входит в головной мозг на уровне моста. Его афференты образуют синапсы в двух ядрах внутри серого вещества моста-спинальном ядре и главном сенсорном ядре. Спинальное ядро тройничного нерва функционально соответствует заднему рогу спинного мозга, тогда как главное сенсорное ядро соответствует ядрам заднего столба спинальных афферентов. Эту аналогию можно распространить и на пути, выходящие из этих ядер. В спинальном ядре афференты от механорецепторов, терморецепторов и болевых рецепторов синаптически переключаются на нейроны, посылающие свои аксоны в ретикулярную формацию и (или) в таламус, куда идут и волокна передне-бокового пучка спинного мозга. В главном сенсорном ядре заканчиваются только афференты от низкопороговых механорецепторов, а аксоны постсинаптических нейронов присоединяются к медиальной петле.
Кроме передачи в таламус, информация, поставляемая афферентами тройничного нерва, используется для двигательных рефлексов мышц области головы. Система тройничного нерва осуществляет жизненно важные функции, особенно у младенцев: например, обеспечивает тактильное узнавание, пищевое поведение и издавание звуков.
Ретикулярная формация.Ретикулярная формация, которая занимает значительную часть ствола мозга, является важным отделом восходящей неспецифической системы (рис. 2-7, Б). Она имеет огромное количество разнообразных афферентных и эфферентных связей, схематически показанных на рис. 2-12, А. Сомато-висцеральные афференты идут по спинно-ретикулярному тракту (передне-бокового пучка) и, возможно, также по проприоспинальным (полисинаптическим) путям, равно как и по соответствующим трактам, выходящим из спинального тройничного ядра. Кроме того, ретикулярная формация получает входные сигналы из всех других афферентных черепных нервов, т. е. от всех органов чувств. Афферентные волокна приходят также из многих других областей головного мозга, например из моторной и сенсорной областей коры больших полушарий, таламуса и гипоталамуса. Эфферентные пути тоже разнообразны: есть спускающиеся в спинной мозг, есть подни-

|
|
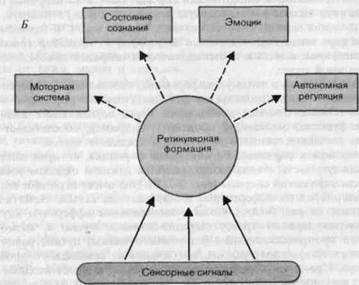
Рис. 2-12. Ретикулярная формация: связи и функции. А. Схема афферентных и эфферентных связей; на рисунке представлена комбинация анатомических и физиологических данных; его назначение-дать представление о большом числе входов и выходов. Б. Наиболее важные функции, приписываемые ретикулярной формации.
мающиеся через неспецифические ядра таламуса к коре, к гипоталамусу и к лимбической системе.
Многообразие афферентных связей проявляется также и на уровне
отдельных нейронов. Микроэлектродные отведения из области ретику
лярной формации показали, что обычно там имеется конвергенция на
один нейрон двух или трех афферентов из числа представленных на
рис. 2-12, А, т.е. npeoбладает полисенсорная конвергенция. Это отличительная черта ретикулярных нейронов. Другие характерные свойства данных нейронов-обширные рецептивные поля, часто представленные билатерально на поверхности тела, большой латентный период реакций на периферическую стимуляцию (связанный с большим числом синапти-ческих переключений), вариабельность реакций (случайные флуктуации числа потенциалов действия, генерируемых в ответ на повторные предъявления стимула, или усиление реакций со временем при действии серий стимулов). Все эти свойства резко отличаются от свойств нейронов специфических ядер, называемых лемнисковыми нейронами соматосенсор-ной системы. Нейроны ретикулярной формации представляют собой пример неспецифических, экстралемнисковых нейронов.
В наших знаниях относительно многообразных функций ретикулярной формации пока еще имеются существенные пробелы. Как схематически представлено на рис. 2-12, Б, полагают, что она вносит вклад в осуществление..ряда функций, которые можно суммировать следующим образом: а) контроль состояния сознания последством влияния на активность кортикальных нейронов и тем самым участие в ритме сон/ бодрствование (ключевой структурой в этом отношении является восходящая ретикулярная активирующая система); б) посредничество_в обес-печении эмоционального влияния сенсорных стимулов, в частности болевых сигналов, передаваемых по передне-боковому пучку путём посылки афферентной информации в лимбическую систему; в) вегетативная регуляция, особенно касающаяся жизненно важных рефлексов (сердечно-сосудистые, дыхательные, глотательные рефлексы, кашель, чихание), когда многие афферентные и эфферентные системы должны работать координированно; г) участие в двигательных механизмах регуляции позы и направленных "движений в качестве важного элемента моторных центров ствола мозга.
В 2.13. Нарисуйте схему поперечного среза спинного мозга, указав афферентные волокна и их связи с сегментарными эфферентами и восходящими трактами, напишите на ней названия.
В 2.14. Предположим, что спинной мозг поврежден: его левая половина полностью перерезана на уровне груди. Какие из перечисляемых ниже симптомов будут наблюдаться?
а) Исчезновение температурной и болевой чувствительности на левой половине тела.
б) Исчезновение температурной и болевой чувствительности в области правой ягодицы.
 68 М. Циммерманн
68 М. Циммерманн
* в) Утрата способности узнавать числа, наносимые на тыльную
поверхность правой стопы. l/г) Утрата способности точно локализовать легкое прикосновение к левой ноге. Верны несколько утверждений. В 2.15. Ретикулярная формация ствола мозга имеет следующие свойства и функции:
а) анатомически она состоит из регулярной сети нейронов с па
раллельными аксонами и дендритами;
б) каждый из ее нейронов получает возбуждение только от
одного органа чувств;
v в) она регулирует общую возбудимость кортикальных нейронов, а тем самым также и состояние сознания;
г) на своем пути в таламус афференты задних столбов синапти-чески переключаются преимущественно в ретикулярной формации.
Дата добавления: 2015-10-05; просмотров: 2166;