Учет с фиксированным уровнем точности и метод обратного биномиального выбора
Проведение учетов численности насекомых – очень важная в практическом отношении, но, как правило, исключительно трудоемкая работа. Как мы уже отмечали, если нужно определить уровень плотности с заданной степенью точности, следует собрать и обработать определенное количество проб. Однако можно не подсчитывать заранее необходимое количество проб, но проводить учеты последовательно, пока полученные результаты не достигнут определенной степени точности. Для такого учета с фиксированным уровнем точности надо на основе предварительных исследовании вычислить так называемую "стоп-линию" и нанести ее на график, где ось ординат – сумма всех последовательно взятых определений плотности (рис.24). Очевидно, что результаты этих последовательных определений будут соответствовать восходящей кривой. Когда эта кривая пересекает "стоп-линию", достигается фиксированный уровень точности.

Рис.24. График последовательного учета численности насекомых с фиксированным уровнем точности. Ось абсцисс – число последовательно берущихся проб, ось ординат – суммарное число всех учтенных насекомых n – число реально выполненных учетов, Q – число обнаруженных во время этих учетов насекомых, n – число 1 учетов, необходимое для определения численности с заданной точностью, М– ожидаемое число обнаруженных насекомых в n пробах (по А.И.Воронцову и др., 1983)
Очевидно, что после проведения некоторого количества учетов мы можем нанести эти данные на график и, наметив примерный ход линии графика, определить общее количество проб, необходимое для данной степени точности учета. На рис. 24 показано, что в результате n 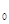 числа проб собрано всего Q насекомых. Продолжая диагональ прямоугольника, доводим ее до пересечения со "стоп-линией". Перпендикуляр, опущенный из точки пересечения M на ось абсцисс, покажет, что необходимо взять
числа проб собрано всего Q насекомых. Продолжая диагональ прямоугольника, доводим ее до пересечения со "стоп-линией". Перпендикуляр, опущенный из точки пересечения M на ось абсцисс, покажет, что необходимо взять 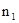 проб.
проб.
Метод обратного биномиального выбора основан на том, что процент проб, содержащих данных насекомых (встречаемость), отражает их реальное количество в природе. Это справедливо, конечно, только при постоянном размере пробы. Расчет основан на формуле отрицательного биномиального распределения. На графике (Рис.25) показано соотношение между долей заселенных деревьев и средним количеством кладок на дерево и доверительные интервалы для двух уровней численности. Такой метод не требует реального подсчета насекомых, и учет на каждом дереве прекращается сразу же после обнаружения хотя бы одной особи. Следовательно, значительно сокращается трудоемкость учета. Этот метод оказывается очень перспективным и в сельскохозяйственной практике.

Рис.25. Линии отрицательного биномиального выборам 90%–ного доверительного интервала при обследовании 50 или 100 деревьев (растений). Ось абсцисс – доля заселенных деревьев, ось ординат – среднее число кладок на одно дерево (по А.И. Воронцову и др., 1983)
Метод последовательного учета
Возможны и другие способы сокращения работы при учете насекомых. Обратим внимание, что при взятии проб мы гарантировали одинаковую ошибку независимо от уровня численности насекомых. Всегда ли в этом есть необходимость? Очевидно, что при практических учетах важно знать не столько реальную численность насекомых, сколько превышает ли она допустимый уровень, возникает ли опасность потери существенной части урожая. Соответственно, при низких уровнях численности можно ее завысить или занизить в несколько раз без какого–либо ущерба. Наоборот, при приближении к критическим уровням точность определения численности приобретает особое значение. Здесь даже небольшая ошибка может привести к существенной потере материальных средств, к ненужному загрязнению местности. План последовательного учета с интервальной оценкой плотности популяции позволяет сократить затраты труда примерно вдвое по сравнению с традиционными методами определения численности. Пример подобного плана последовательного учета приведен на рисунке 26. Здесь показаны три градации: сильное, среднее и слабое заражение вредителем. Для практических нужд, по–видимому, достаточно только двух градаций: выше порога вредоносности и ниже него.
Эти планы представляют собой оси координат. Ось абсцисс – последовательность проб, которые берутся для учета (1,2,3 и т.д.), а ось ординат – сумма учтенных насекомых, обнаруженных в этих пробах. На осях ординат нанесены также зоны неопределенности, каждая из которых ограничена двумя параллельными линиями, идущими под углом и симметрично по отношению к нулевой точке. На рисунке показано по две такие зоны, но при расчете, исходя из порога вредоносности, достаточно одной. Для расчета этих зон, точнее линий, их ограничивающих, необходимо на основе предварительных учетов найти среднее число особей на пробу и дисперсию этих данных, выбрать порог вредоносности, а также уровень достоверности нашего учета.
Когда эти зоны построены, мы начинаем наносить на график результаты последовательно взятых проб. Получающаяся восходящая кривая в зависимости от ее наклона продолжается за пределами зоны неопределенности выше, или ниже ее. Соответственно, численность учитываемых насекомых оказывается выше или ниже порога вредоносности.

Рис.26. План последовательного учета яиц шелкопряда–монашенки Porthetria monacha L. Ось абсцисс – число последовательно берущихся проб (по 100 г хвои), ось ординат – суммарное количество обнаруженных яиц. Пунктирной линией показаны результаты реального учета, при сильной зараженности деревьев монашенкой (по А.И.Воронцову и др., 1983)
Более подробный расчет для последовательного учета приведен в статье А.И.Воронцова и др.(1983).
Метод корреляционных функций
Регулярные обследования с целью учетов насекомых проводятся многими организациями как в сельском, так и в лесном хозяйстве. Естественная цель этих обследований – во-время заметить начинающееся массовое размножение вредного вида. Однако массовые размножения редко ограничены каким-то одним полем или лесом. Изменения метеорологических условий, благоприятные для массового размножения сразу охватывают большой географический район. Кроме того, насекомые активно мигрируют, что приводит к определенному выравниванию плотностей популяций. Поэтому во многих случаях число учетов в районе можно сократить.
Ф.Н.Семевский предложил для рационализации системы учетов так называемый метод корреляционных функций. Суть его сводится к следующему. Определяют плотность популяций какого-либо вида во многих парах равноудаленных точек, например, на расстоянии 1 км друг от друга. Вполне вероятно, что если в одной точке из такой пары наблюдается высокая зараженность, то в другой на расстоянии в 1 км она тоже не будет низкой. И, наоборот, если вредитель в одной точке совсем отсутствует, то на расстоянии в 1 км его плотность навряд ли будет высокой. Следующие шаги – определение корреляции для пар точек, удаленных друг от друга на 10, 100 км и т.д. Очевидно, что чем дальше удалены друг от друга точки, тем менее сходны плотности популяций в этих точках. Возможен и такой случай, когда корреляция плотностей исчезает даже на близком расстоянии.
Если отложить на графике коэффициенты корреляции плотностей для все более удаленных точек мы получим график корреляционной функции, который может быть взят за основу для рационализации процесса учета в пределах данного географического района.
Методы учета относительной численности
Все перечисленные нами выше методы учета были направлены на то, чтобы получить представление о реальном уровне численности объекта или, по крайней мере не превышает ли эта численность какую-то ранее установленную нами границу. Однако существует ряд методов, с помощью которых можно судить о численности лишь косвенно, либо по какой–то части популяции, либо по результатам деятельности насекомых. Такие методы дают ориентировочное представление о численности насекомых и о тенденциях ее изменений.
В качестве примера М.С.Гиляров(1975) приводит учет почвенной мезофауны наблюдателем, идущим за плугом. Конечно, есть лишь определенная корреляция между количеством личинок насекомых, обнаруженных этим способом, и абсолютной численностью.
Строго говоря, и метод энтомологического кошения также дает лишь ориентировочную оценку численности, равно как и определение плотности по числу поврежденных растений или листьев, цветов, бутонов, плодов, а также по числу экскрементов насекомых или их линочных шкурок.
Очень часто о численности популяции судят по результатам уловов насекомых с помощью различных ловушек. Все эти ловушки можно разделить на две категории: без привлекающих агентов, в которые насекомые попадают случайно, и специально привлекающие насекомых. Первые всегда собирают значительно меньше насекомых, чем вторые. Однако результаты, полученные с помощью ловушек без привлекающих агентов гораздо легче интерпретируются. Дело в том, что привлекательность факторов, например, света или химических стимулов, в значительной степени зависит от погоды и физиологического состояния насекомых. Поэтому малый сбор, полученный с помощью такой ловушки, можно трактовать либо как реальную низкую численность объекта, либо как снижение привлекательности фактора при той же или даже большей численности.
Среди ловушек без привлекающих факторов следует упомянуть земляные ловушки Барбера (банки, вкопанные в почву), оконные ловушки, где случайно пролетающие насекомые ударяются о стекло и падают в фиксирующий раствор (М.Н.Самков, Д.Б.Чернышев, 1983), сачки, закрепленные на общей вращающейся оси, ловушки, всасывающие насекомых вместе с воздухом, ловушки Малеза, представляющие собой куполообразный тент, открытая вершина которого является вершей с фиксирующим раствором. Все эти ловушки рассчитаны на случайное попадание в них передвигающихся насекомых. Однако величина их улова зависит не только от численности популяций насекомых, но и от уровня подвижности. Кроме того, в зависимости от времени суток и года, от погоды или по другим причинам пути и уровни полета насекомых могут меняться. Достаточно лишь насекомым лететь немного выше, чтобы не попадать ни во всасывающие воздух, ни в оконные ловушки.
Ловушки с привлекающими насекомых факторами также очень разнообразны. Простейшие из них – пучки выполотой травы и сорняков для привлечения гусениц подгрызающих совок, а также жуков–щелкунов и жужелиц. В ряде случаев для увеличения сбора ловушками Барбера также применяют различные, обычно пищевые приманки. На практике широко используют целый набор ловушек с половыми феромонами, а также светоловушки.
Совершенно очевидно, что ни одна из этих ловушек не может дать надежной информации ни о времени появления тех или иных насекомых, ни об их реальной численности. В частности, фенологические сроки появления имаго ряда бабочек и жуков, полученные с помощью светоловушки, могут отставать от реальных на 10–15 дней. Причина этого – изменение времени лета имаго по отношению к световому дню. Так, в мае лет может завершаться еще до наступления темноты или по крайней мере еще до достижения того уровня естественной освещенности, когда становится возможным лет насекомых данного вида на свет.
Большое количество насекомых, выловленных светоловушкой, не всегда означает, что реальная численность местной популяции очень высока. Некоторые бабочки могут привлекаться светом во время их массовых миграций, так сказать – "на пролете". Тем не менее сбор насекомых с помощью феромонных и световых ловушек широко применяется, благодаря простоте их использования и получению большого количества материала. Вопрос же об истинной ценности этих материалов, как правило, не рассматривается.
4. ВОЗРАСТНАЯ И ПОЛОВАЯ СТРУКТУРА ПОПУЛЯЦИИ
Возрастная и половая структура популяции насекомых – важнейшие показатели состояния популяции, подъема или спада ее численности. Анализ гибели насекомых на разных стадиях развития (таблицы выживания) является основой для понимания динамики численности популяций. Во многих случаях картина возрастного и полового состава популяций принципиально отличается от таковой у позвоночных, описанной в многочисленных руководствах по экологии. У общественных насекомых, помимо возрастной и половой структуры популяций, имеет место также разделение всех особей на касты, которое мы здесь не рассматриваем.
Возрастной состав популяции
Под возрастным составом популяции насекомых следует понимать соотношение численностей насекомых на разных стадиях развития или имаго разных возрастов в каждый данный момент времени. Если неблагоприятный сезон могут пережить только насекомые на одной какой–либо стадии, то именно они и будут представлены в течение всего этого сезона. Наличие в это время только половозрелых особей не будет означать, что популяция вымирает, равно как наличие только яиц не свидетельствует о бурном развитии популяции. Однако и в дальнейшем популяция в каждый момент времени может быть представлена только одной или двумя стадиями развития. Это особенно типично для моновольтинных видов. Чем больше поколений проходит за год, тем большее количество стадий можно найти одновременно.
Поэтому широко известные возрастные "пирамиды", показывающие соотношение особей разных возрастов у позвоночных и дающие возможность оценить способность популяции к увеличению численности, для насекомых могут быть получены лишь в редких случаях. Однако, например, в случае длительного выращивания большого количества особей в лабораторной культуре соотношение их численностей на разных стадиях развития является хорошим показателем состояния культуры. Для периода экспоненциального роста численности и начального освоения субстрата типично большое количество личинок. Наоборот, повышение численности личинок старших возрастов и имаго соответствует прекращению нарастания численности насекомых в культуре и исчерпанию жизненных ресурсов (Никитина–Петрова и др., 1986).
Таблицы выживания
Насекомые, подобно большинству других беспозвоночных, отличаются низкой выживаемостью, т.е. малой вероятностью выживания особи в процессе развития. Действительно, численность популяции в природе при неизменных средних условиях всегда остается примерно на одном и том же среднем уровне, а плодовитость же одной самки насекомых колеблется от нескольких десятков до нескольких тысяч яиц. Это означает, что шанс каждой особи дойти до половозрелого состояния очень мал. Для насекомых, в общем, типична высокая смертность на ранних стадиях развития. Особенно часто погибают яйца и личинки младших возрастов. Соответственно, кривая выживания насекомых в большинстве случаев вогнутая, что показывает сначала быстрый, а потом замедленный спад численности по мере развития.
Анализ выживания насекомых определенного вида на разных стадиях их развития может быть представлен как таблица выживания. Составление этих таблиц – удобный способ проследить изменения возрастного состава популяции на протяжении развития и, что особенно важно, проанализировать конкретное влияние различных биотических и абиотических факторов на численность популяции.
Остановимся сначала на основных принципах составления таблиц выживания, а затем рассмотрим их конкретные примеры. В самом простом случае мы имеем дело с моновольтинным видом и неперекрывающимися поколениями. Предположим, что плотность популяции только что отложенных яиц (а 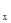 ) равна 1000 на один м
) равна 1000 на один м 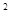 . На протяжении развития 900 яиц погибает по разным причинам, которые можно учесть отдельно (например, от яйцевых паразитов, вымерзания, вымачивания и т.д.). Тогда плотность популяции личинок первого возраста (а +1) будет равна 100 на м . Предположим далее, что позже можно обнаружить только 10 личинок второго возраста на 1 м (а +2). Следовательно, ряд, показывающий плотность популяции на этих трех разных стадиях будет следующим: 1000, 100 и 10.
. На протяжении развития 900 яиц погибает по разным причинам, которые можно учесть отдельно (например, от яйцевых паразитов, вымерзания, вымачивания и т.д.). Тогда плотность популяции личинок первого возраста (а +1) будет равна 100 на м . Предположим далее, что позже можно обнаружить только 10 личинок второго возраста на 1 м (а +2). Следовательно, ряд, показывающий плотность популяции на этих трех разных стадиях будет следующим: 1000, 100 и 10.
Если принять исходную плотность за единицу, то доля выведшихся личинок первого возраста (I +1), будет равна 0,1, а личинок второго возраста (I +2) – 0,01. Соответственно, смертность (d ) на стадии яйца равна 1 – 0,1= 0,9 , а личинок первого возраста 0,1 – 0,01= 0,09.
Важной характеристикой является также коэффициент смертности – q . Он равен частному от деления d /I , т.е. соотношению доли погибших особей к доле выживших. В нашем случае он равен 0,9/1,0 = 0,9 для яиц и 0,09/0,1=0,9 для личинок первого возраста.
При расчетах удобнее оперировать не с самими числами, а с их логарифмами. Это существенно, в частности, при расчете интенсивности смертности (k ), которая представляет собой разность log a (логарифм числа особей в начале развития стадии) и log a +1 (логарифм числа особей в конце развития этой – начале развития следующей стадии). В нашем случае данный коэффициент для стадии яйца равен 3–2=1 (log 1000=3; log100=2). Ту же величину мы получим для личинок первого возраста (2–1=1). Оба эти показателя – коэффициент смертности и интенсивность смертности имеют большое значение при анализе динамики численности популяции. Анализируя таблицы выживания за ряд поколений, можно проследить, как численность популяции связана с уровнем того или иного фактора среды.
В качестве одного из конкретных примеров рассмотрим таблицу выживания бабочки зимней пяденицы Operophtera brumata L., опубликованную в книге Д.К.Варли, Д.К.Градуелла и М.П.Хассела (1978) (табл.2).
Таблица 2
Таблица выживания для зимней пяденицы (на 1955–1956 г.) (по Д.К.Варли, Д.Р.Градуеллу и М.П.Хасселу, 1978 с изменениями)
| Показатели | a
| m
| d
| log a
| k
|
| Имаго | 4,4* | ||||
| Яйца | 658,0 | 2,82 | |||
| Гусеницы последнего возраста | 96,4* | 561,6 | 0,85 | 1,98 | 0,84 |
| зараженные Cyzenis | 90,2 | 6,2* | 0,06 | 1,95 | 0,03 |
| другими паразитами | 87,6 | 2,6* | 0,02 | 1,94 | 0,01 |
| микроспоридиями | 83,0 | 4,6* | 0,04 | 1,92 | 0,02 |
| Куколки | |||||
| съеденные хищниками | 28,4 | 54,6 | 0,66 | 1,45 | 0,47 |
| зараженные паразитами | 15,0 | 13,4* | 0,46** | 1,18 | 0,27 |
| Имаго – 1956г. | 7,5* |
* звездочкой обозначены фактические наблюдения, остальные величины – производные от них.
** смертность от хищников и паразитов рассмотрена как последовательные явления.
a– число особей, оставшихся в живых (на 1 м  ); m– число погибших на данной стадии особей (на 1 м ); d– смертность (доля особей, погибших на данной стадии отданной причины); k– интенсивность смертности (log a– log a+1).
); m– число погибших на данной стадии особей (на 1 м ); d– смертность (доля особей, погибших на данной стадии отданной причины); k– интенсивность смертности (log a– log a+1).
Бабочки этого вида выводятся из куколок поздней осенью. Практически бескрылые самки поднимаются по стволам деревьев вверх и откладывают яйца в трещинах коры. Чтобы определить численность бабочек, на стволах деревьев закрепляли вершеобразные ловушки. Эти ловушки располагали на 5 деревьях с общей площадью проекции полога на землю 282м , по две на каждое дерево. Каждая из ловушек охватывала 1/8 периметра ствола, поэтому улов всеми ловушками соответствовал 1/4 численности всех самок на этом участке. Всего было поймано 413 самок. Очевидно, что на участке их должно было быть 413х4=1652, т.е. около 5,85 на 1 м . Одна четверть самок погибла в ловушках, следовательно, плотность популяции самок, отложивших яйца в кронах деревьев, была равна 5,85x0,75 = 4,39.
По вскрытиям было установлено, что самки в среднем откладывали по 150 яиц. Следовательно в расчете на один м было отложено около 658 яиц. Гусеницы, готовые к окукливанию, спускались вниз на шелковинках. 5ы определить их численность, под каждое из пяти деревьев ставили І два лотка. В лотках накапливалась дождевая вода, в которой тонули спускающиеся гусеницы. Площадь каждого лотка была равна 0,5 м . Таким образом, можно было подсчитать число уходящих на окукливание гусениц на всем участке, а, кроме того, вскрывая пойманных гусениц, определить их зараженность мухой-тахиной Cyzenis albicans Fll., другими паразитами и микроорганизмами. Если на 1 м приходилось 658 яиц, то окукливающихся гусениц было всего 96,4, т.е. смертность достигала 0,85. Причины этой смертности остались неясными. Во всяком случае, из большинства яиц выводились гусеницы, а гибель от птиц, скорее всего, была очень незначительной. Возможно, что здесь имела место массовая эмиграция гусениц с деревьев, запаздывающих с весенним распусканием почек.
Предполагалось, что все гусеницы, достигшие поверхности почвы, окукливались. Выживание куколок можно было определить по количеству бабочек, пойманных тем же методом следующей осенью. Для подсчета паразитов куколок землю под деревьями накрывали теми же самыми лотками, которые использовали ранее для сбора спускающихся для окукливания гусениц, перевернув их и воткнув краями в почву. В углы лотков были вмонтированы стеклянные пробирки. Паразиты, выходя из почвы под лотком, стремились к свету и скапливались в этих пробирках.
Интересна также приводящаяся теми же авторами таблица выживания паразитической тахины Cyzenis (табл.3). Громадная гибель яиц этой мухи явно связана с тем, что они откладываются не на хозяина, а на листья, поврежденные гусеницами, и шанс для яиц быть проглоченными гусеницами очень мал.
Естественно, что в зависимости от конкретных условий и географического района таблицы выживания одного и того же вида могут существенно различаться. Выживание также будет различным и при разных уровнях численности. Однако на примере сосновой пяденицы Bupalus piniarius L. было показано, что вспышка массового размножения этого вида, хотя и меняет количественные соотношения, не приводит к качественным изменениям таблицы выживания (N.Broekhuizen et al., 1994).
Таблица 3
Таблица выживания для мухи-тахины Cyzenis (по Д.К.Варли, Д.Р.Градуеллу и М.П.Хасселлу 1978, с изменениями)
| a
| log a
| k
| |
| Имаго – 1955 г. | 0,15 | ||
| Потенциальное число яиц | 300,0 | 2,48 | |
| Личинки в гусеницах | 6,2 | 0,79 | 1,69 |
| Имаго – 1956 г. | 0,3 | -1,48 | 1,31 |
Примечание: обозначение граф те же, что и на таблице 2.
Дата добавления: 2015-05-16; просмотров: 1047;