Фазовый портрет динамики численности 1 страница
Динамику численности популяции любого насекомого можно представить в виде так называемого фазового портрета (рис.37 – А.С.Исаев и др., 1984). Независимая переменная x на этом графике – плотность популяции (ось абсцисс), зависимая у – коэффициент размножения (ось ординат). Очевидно, что при у=1 популяция стабильна, если у>1, то популяция возрастает, а у<1 – убывает. Согласно сказанному выше, коэффициент размножения обратно пропорционален плотности популяции. Поэтому мы вправе представить зависимость у от x в виде гиперболы, кривой, соответствующей обратной пропорциональности (y=а/x).
Проведем на графике прямую у=1. Точка пересечения этой прямой с гиперболой (а) покажет устойчивое состояние популяции. При вызванном какими–либо внешними обстоятельствами (для фитофага обычно ослаблением кормового растения) увеличении плотности популяции коэффициент размножения снизится и популяция автоматически вернется к прежнему устойчивому уровню. При снижении численности популяции, наоборот, будет иметь место увеличение коэффициента размножения и возврат к той же устойчивой точке.
 Рис.37. Фазовый портрет динамики численности популяции фитофага (схема – ось абсцисс – плотность популяции, ось ординат – коэффициент размножения): I– область действия регуляторных механизмов со слабой инерцией (ограничена кривыми у' и у
Рис.37. Фазовый портрет динамики численности популяции фитофага (схема – ось абсцисс – плотность популяции, ось ординат – коэффициент размножения): I– область действия регуляторных механизмов со слабой инерцией (ограничена кривыми у' и у 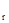 ); II – область максимальной инерционности регуляторных механизмов (ограничена кривыми у и у
); II – область максимальной инерционности регуляторных механизмов (ограничена кривыми у и у 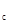 ); III – область действия безынерционных механизмов регуляции (ограничена кривыми y и y'). Кривые: 1 – нижняя с граница фазового портрета у', 2 – верхняя граница фазового портрета у ", 3 – пороговая у , 4 – буферная у
); III – область действия безынерционных механизмов регуляции (ограничена кривыми y и y'). Кривые: 1 – нижняя с граница фазового портрета у', 2 – верхняя граница фазового портрета у ", 3 – пороговая у , 4 – буферная у 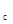 , 5 – статическая y
, 5 – статическая y  . Фазовая траектория вспышки: bc – фаза нарастания численности популяции, cd – максимума, de – разреживания, eh – депрессии, ha – фаза восстановления стабильной численности. Характерные точки: x
. Фазовая траектория вспышки: bc – фаза нарастания численности популяции, cd – максимума, de – разреживания, eh – депрессии, ha – фаза восстановления стабильной численности. Характерные точки: x 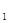 – значение стабильной плотности популяции, x – пороговая плотность, x – оптимальная плотность, x
– значение стабильной плотности популяции, x – пороговая плотность, x – оптимальная плотность, x 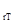 – минимально предельная плотность на фазе максимума вспышки, x
– минимально предельная плотность на фазе максимума вспышки, x 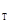 – максимально предельная плотность (по А.С. Исаеву и др., 1984)
– максимально предельная плотность (по А.С. Исаеву и др., 1984)
Однако сказанное справедливо, если действуют только безынерционные механизмы, регулирующие численность в пределах одного поколения. В природе же на этом уровне численности обычно основными являются запаздывающие с действием инерционные механизмы (например, численная реакция энтомофага или изменения биохимических свойств растения под влиянием интенсивного питания фитофага). Благодаря такой инерционности популяция "ускользает" от сдерживающих ее факторов и переходит к массовому размножению. Сначала увеличиваются и коэффициент размножения, и численность популяции (отрезок кривой Ь – с). Точка с соответствует предельному процветанию популяции, когда высокая плотность сочетается с максимальным коэффициентом размножения. Однако начиная с этой точки вступают в действие безынерционные механизмы (исчерпание ресурсов, эпизоотии). Поэтому коэффициент размножения постепенно снижается. Численность же продолжает нарастать до тех пор, пока этот коэффициент остается больше единицы. Далее же и численность начинает убывать. Кривая переходит в зону действия инерционных механизмов с точки d . Уже размножившиеся с начала вспышки энтомофаги еще более снижают и коэффициент размножения, и соответственно численность. Впрочем, снижение численности после вспышки может быть связано с интенсивной эмиграцией (М.Д. Корзухин, Ф.Н. Семевский, 1992).
Далее с понижением плотности пресс регулирующих факторов ослабевает. Начиная с точки Л коэффициент размножения возрастает, но численность популяции продолжает снижаться, поскольку он еще меньше единицы.
Помимо уже известной нам основной параболы на график нанесены также кривые, ограничивающие пределы возможного для данного вида фазового портрета и связанные с его биологическими свойствами и возможностями биоценоза: нижняя граница фазового портрета – у' и верхняя граница – у". Выход системы за верхнюю границу возможен лишь на очень короткое время, которое значительно меньше времени генерации, так как мощные безынерционные процессы сразу же возвращают систему в ее пределы. Нижние пределы менее определенны, популяция может вымирать и восстанавливаться заново за счет мш–рантов. На график еще нанесены две кривые 3 и 4 (у и y ), которые показывают область максимального воздействия инерционных процессов.

Рис. 38. Фазовый портрет динамики численности сибирского шелкопряда Dendrolimus sibiricus Tshtvr. Оси абсцисс и ординат представлены в логарифмах, цифры у кривых обозначают годы, наклонной прямой показаны пороговые значения соотношения плотности популяции и коэффициента ее размножения, приводящие к вспышке, у 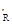 – значение коэффициента размножения, приводящее к вспышке при стабильной в норме численности x
– значение коэффициента размножения, приводящее к вспышке при стабильной в норме численности x 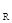 . Прочие обозначения те же, что на рис. 37 (по А.С.Исаеву и др., 1984)
. Прочие обозначения те же, что на рис. 37 (по А.С.Исаеву и др., 1984)
Таким образом, благодаря инерционности регулирующих популяцию факторов, вместо немедленного возврата к устойчивому состоянию возникает определенный циклический процесс, который в конце концов тоже приводит популяцию к устойчивому состоянию. Именно такие циклические процессы обнаруживаются при длительном наблюдении за численностью многих насекомых. Так, на рис.38 показан фазовый портрет динамики численности сибирского шелкопряда (А.С.Исаев и др., 1984) за срок с 1956 по 1972 г. Здесь четко выявляются два таких популяционных цикла и зона наибольшей стабильности (поскольку на этом рисунке применена логарифмическая шкала, то коэффициент размножения, равный единице, представлен как lg 1=0).
Очевидно, легче всего искусственно регулировать численность на отрезке a–b, когда создаются благоприятные условия для фитофага и пресса природных энтомофагов оказывается еще недостаточно для сдерживания его размножения. На этом этапе выпуск небольшого количества искусственно разведенных энтомофагов может снизить еще небольшую численность фитофага до стабильного уровня. Гораздо больше усилий понадобится, чтобы стабилизировать численность на фазе b–с, когда равновесие уже нарушено. Начиная с точки с и далее любые меры искусственного регулирования численности, за исключением самых радикальных и опасных для биоценоза, могут привести к продлению вспышки массового размножения. К сожалению, именно на этой фазе c–d при самой высокой численности массовое размножение становится очевидным, численность переходит экономический порог вредоносности и надо принимать срочные меры.
Весь цикл, изображенный на фазовом портрете, у лесных насекомых занимает несколько лет. В сельскохозяйственной же практике, особенно у поливольтинных насекомых, все фазы этого цикла протекают значительно быстрее. Во многих случаях из–за общей неустойчивости агроценоза популяция выходит из под контроля естественных сдерживающих факторов практически ежегодно. В некоторых же случаях пресс естественных врагов оказывается настолько слабым, что вспышка принимает перманентный характер.
Принцип ультрастабильности
Воздействие любого фактора на популяцию определяется уровнем численности последней. При этом каждый из факторов оказывает максимальное воздействие на определенном уровне плотности, а при более высоких уровнях его значение может быть невелико. Поэтому с ростом численности популяции то одни, то другие факторы оказываются регулирующими ее численность и являются преградой на пути массового размножения (рис.39).
Такая ступенчатая схема регуляции обеспечивает ее ультрастабильность (Г.А.Викторов, 1967). На наиболее низком уровне численности популяция сдерживается многоядными хищниками и паразитами. На более высоком уровне основным регулятором являются специализированные энтомофаги. Эпизоотии и нехватка корма реально влияют на численность при ее высоком уровне. В природе, однако, достижению такой численности препятствует массовая эмиграция или дифференциация популяции во времени, например по длительности диапаузы.
Диапазон плотности, в котором фактор может регулировать численность популяции, связан прежде всего со способностью этого фактора быстро реагировать на изменение плотности, с его инерцией. Так, увеличение численности жертвы может приводить к увеличению численности энтомофага только в его следующем поколении, т.е. инерция энтомофагов как фактора очень велика. Меньше инерционность у возбудителей заболеваний, хотя зараженность обычно возрастает не сразу, а часто на протяжении нескольких поколений. Инерция здесь может уменьшаться за счет повышения восприимчивости, связанного, например, с ухудшением или недостатком корма или со слишком холодной и сырой погодой. Внутривидовые регуляторные механизмы действуют незамедлительно, и их эффект не связан с численностью предыдущих поколений (конкуренция за пищу и пространство, каннибализм). По-видимому, прежде всего при высоких уровнях численности должно проявляться регулирующее действие абиотических факторов, которые также отличаются быстрым действием).
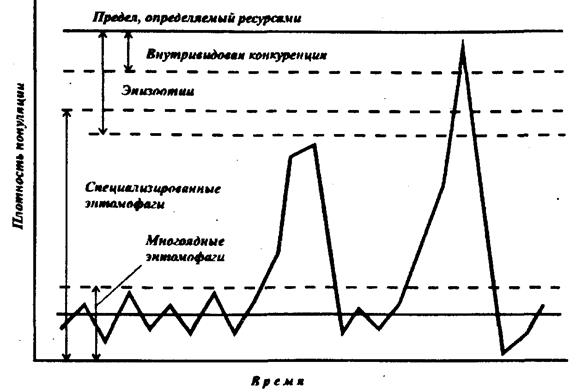
Рис.39. Схема порогов и зон активности основных механизмов регуляции численности насекомых (по Г.А.Викторову, 1967)
Таким образом, на низких уровнях численности регуляторами являются: прежде всего факторы с большой инерцией, а на высоких – быстродействующие.
Наличие инерционных механизмов делает всю систему менее жесткой. Примечательно, что у видов, способных к массовым размножениям, например у сибирского шелкопряда, безынерционные механизмы включаются только на довольно высоких уровнях численности. Паразиты же при создавшихся благоприятных для размножения этого вида условиях часто не "успевают" за ростом численности хозяина. У видов с более устойчивым уровнем популяции инерционность в регуляции невелика, например, их специализированные паразиты способны к очень быстрому размножению.
Очевидно, что при попытках увеличить численность каких–либо полезных видов, исходно уже обитающих в данном биотопе, за счет выпуска специально разведенных в лаборатории насекомых немедленно включаются мощные регулирующие механизмы, которые быстро доводят эту численность до исходного уровня. Так, даже после многократных выпусков трихограммы ее численность в поле быстро снижается до минимума. То же самое происходит при промышленном разведении пчел–мегахил, которые при выставлении искусственных гнездовий в поле в массе заражаются паразитами (В.И. Голиков, 1989).
Наоборот, для сельскохозяйственных растений особую опасность представляют насекомые–фитофаги, лишь недавно обосновавшиеся в данном районе (адвентивные виды), поскольку их биоценотические отношения с местными видами еще не сложились и сдерживающее влияние хищников и паразитов почти отсутствует. В таких случаях оказывается особенно эффективной интродукция энтомофагов из тех районов, откуда происходит данный вид фитофага (С.С. Ижевский, 1983).
Типы динамики численности
На примере лесных насекомых можно различить три основных типа динамики численности: стабильный, продромальный и эруптивный (А.С. Исаев и др., 1984).
У видов со стабильной численностью имеют место лишь небольшие колебания численности в зоне точки устойчивости. Это типичные К–стратеги с низкой миграционной активностью и с малой плодовитостью. Фитофаги с подобным типом динамики численности никогда не наносят существенного вреда растению, на котором они питаются. Такие виды составляют около 50–60% общего видового состава данной местности.
В случае продромального типа динамики численности благоприятные условия (погода, наличие ослабленных растений и т.д.) приводят к существенному нарастанию численности в отдельные годы. Типичные представители: бабочка–траурница, дубовый коконопряд, большинство насекомых – ксилофагов (короеды, усачи и другие). Массовое размножение этих видов может быть описано тремя фазами: нарастание, максимум и разреживание. Депрессия численности, как правило, отсутствует.
Наконец, эруптивные виды способны на длительный срок отрываться от контроля энтомофагов и давать вспышки массового размножения. По–видимому, все они r–стратеги, отличающиеся высокой плодовитостью и миграционными способностями. Примерами таких насекомых являются сибирский и непарный шелкопряды. Цикл динамики численности этих видов складывается из пяти фаз: нарастание, максимум, разреживание, депрессия и восстановление. Вспышка размножения может возникать одновременно в пределах всей ландшафтной зоны или же сначала в наиболее благоприятной зоне с последующим ее распространением. Возможны и локальные вспышки в ослабленных по тем или иным причинам лесах. Вспышки могут стать и панзональными (пандемическими), охватывая при этом несколько ландшафтно–географических зон.
Модели динамики численности
Моделирование процесса – это единственно возможный путь к его пониманию. В экологии насекомых часто разрабатываются словесные модели, более перспективно математическое моделирование, использующее не только качественную, но и количественную информацию о процессах.
Иногда применяют и другие способы моделирования (например, известный эксперимент Холлинга, в котором вслепую искали на столе диски из наждачной бумаги). В настоящем разделе мы остановимся только на некоторых основных принципах, используемых при разработке математических моделей динамики численности.
Известно три класса моделей, описывающих динамику численности популяций: 1) эмпирико–статистические; 2) элементарных экологических процессов (субмодели) и 3) жизненных систем популяции (имитационные модели).
Эмпирико–статистические модели основаны на статистическом анализе корреляций между реальной плотностью популяции изучаемого вида и различными факторами среды. Эти модели могут быть очень полезными для приблизительного прогноза численности. Влияние каждого фактора представляется обычно отдельными коэффициентами полинома – уравнения многофакторной регрессии типа
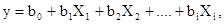
где Х –X 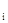 – значения каждого из факторов, b
– значения каждого из факторов, b 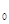 –b – значения коэффициентов регрессии. Зависимости для каждого фактора могут быть не только линейными, но сколь угодно сложными. Очевидно, что сами коэффициенты регрессии обычно не имеют четкого биологического смысла, и в зависимости от ситуации, т.е. уровня факторов и их взаимодействий, могут меняться. В качестве факторов обычно выбирают плотность популяции предшествующего поколения и различные метеорологические показатели.
–b – значения коэффициентов регрессии. Зависимости для каждого фактора могут быть не только линейными, но сколь угодно сложными. Очевидно, что сами коэффициенты регрессии обычно не имеют четкого биологического смысла, и в зависимости от ситуации, т.е. уровня факторов и их взаимодействий, могут меняться. В качестве факторов обычно выбирают плотность популяции предшествующего поколения и различные метеорологические показатели.
Некоторые модели элементарных экологических процессов, например, внутривидовой конкуренции, взаимодействия паразит–хозяин были рассмотрены нами в предыдущих разделах. Эти модели были направлены не на анализ конкретных данных, а на понимание сущности процесса. Как было также показано выше, они могут иметь и некоторую прагматическую ценность.
Модели жизненных схем популяций, совмещающие в себе несколько моделей элементарных процессов, оказываются наиболее ценными как для понимания процессов, протекающих в природе, так и для прогнозирования. Кроме того, эти модели могут быть основой для управления экологическими процессами.
Субмодели соединяются воедино соответственно формуле
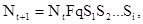
где N 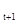 – численность популяции в последующем поколении, N
– численность популяции в последующем поколении, N 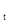 – численность предыдущего поколения, F – плодовитость, q – доля самок в предыдущем поколении, S -S – выживаемость особей по отношению к тому или иному фактору.
– численность предыдущего поколения, F – плодовитость, q – доля самок в предыдущем поколении, S -S – выживаемость особей по отношению к тому или иному фактору.
Помимо субмоделей в число этих показателей могут быть включены и коэффициенты, полученные методом статистической регрессии. Исходным материалом для таких моделей служат таблицы выживания, по возможности многолетние. Таблицы выживания необходимо составлять не только для изучаемого вида, но и для его паразитов и хищников, а также кормового растения.
Так как и вариации абиотических факторов, и биологические взаимодействия не могут быть полностью предсказаны, то желательно, чтобы и модель не была жестко детерминирована. Она должна включать в себя дополнительные элементы случайного характера и, соответственно, давать прогноз с той или иной степенью вероятности.
Вопросы моделирования экологических процессов более подробно изложены во многих монографиях, например книгах Ф.Н.Семевского и С.М.Семенова (1984), а также М.Д.Корзухина и Ф.Н.Семевского (1992).
Модели численности, построенные по этим принципам, имеют большое значение как для практики, так и для экологии в целом. Однако реальная ситуация, складывающаяся в биоценозе, всегда намного сложнее, чем ее отражение в модели. Дело не только в большом количестве факторов и их взаимодействий. Как правило, если судить по таблицам выживания, многие из них оказываются несущественными. Стремление охватить возможно большее количество факторов обычно приводит к неоправданному усложнению модели. Кроме того, согласно экологическому закону минимума влияние фактора оказывается тем сильнее, чем дальше его величина от оптимума. Соответственно, если учеты численности проводились в то время, когда фактор находился на оптимальном уровне, то его влияние останется незаметным и он будет исключен из анализа как несущественный. Однако достаточно относительно небольшого отклонения этого фактора от оптимума, чтобы исказить построенную нами картину явления. Такое несовершенство всех моделей сложных экологических явлений может проявиться как при изменениях во времени (например, от года к году), так и в пространстве (от одного биотопа к другому), поскольку и учеты численности, и составленные на их основе таблицы выживания опираются на результаты наблюдений за данной популяцией и в данное время. Поэтому модель справедлива лишь при изменениях всех факторов в пределах, которые имели место во время наблюдений.
При построении моделей возникают и другие сложности. Очевидно, что точность измерений (определения численности в природе) не очень велика, а объединение неточных данных еще больше увеличивает вероятность ошибки. Далее, плодовитость часто принимают за постоянную величину. На деле же абиотические факторы в их обычных пределах не столько вызывают смертность, сколько влияют на плодовитость. Кроме того, генетическая структура популяции может спонтанно меняться во времени, что должно отражаться как на плодовитости, так и на устойчивости насекомого по отношению к неблагоприятным факторам среды и заболеваниям.
Исключительно важно учитывать пространственную разнокачественность той местности, где проводятся наблюдения и миграции насекомых. Условия могут очень существенно варьировать даже в пределах одного биотопа. Напомним, что только температура в пределах одного растения может различаться в пределах 15 и более градусов. В разные годы могут оказаться наиболее пригодными для жизни насекомых данного вида то одни, то другие участки микрорельефа. Чем разнообразнее условия в пределах биотопа, чем больше шансов имеет популяция для выживания.
Миграции имеют очень большое значение в жизни насекомых, но еще недостаточно изучены. Дальние миграции относительно крупных насекомых по направленности и расстояниям могут быть сравнимы с перелетами птиц. Пассивные же миграции мелких насекомых вместе с токами воздуха фантастичны по масштабам вовлечения в них насекомых и также достигают по дальности сотен, а иногда и тысяч километров. Роль миграций обычно недооценивают, потому что очень трудно определить их количественные масштабы. Даже если это миграция на небольшие расстояния, она приводит не только к изменениям численности локальной популяции, но и к обмену генетической информацией и изменениям генетической структуры популяции.
Только в последнее время на основе современной компьютерной техники начали появляться модели, учитывающие жизнь популяции в пространстве. Такая модель была составлена для хлопковой совки Helicoverра armigera (Hubner) и близкого к ней вида H.punctigera Wallengren. (M.L.Dillon, G.P.Fitt, 1991).O6a эти вида являются вредителями хлопчатника в Австралии.
Основным отличием этой модели является введенная в компьютер карта местности, разделенная на относительно небольшие ячейки, характеризующиеся своим растительным покровом и почвенно–климатическими условиями. В компьютер вводится отдельно для каждого такого участка информация о состоянии и фазе развития основного растения, о численности и развитии этих совок и метеорологические условия. В модель включены имитация развития яиц, гусениц и куколок, миграции имаго в зависимости от их физиологического состояния, температуры, направления и скорости ветра, а также откладки яиц в зависимости от привлекательности растений данного вида и на данной стадии их развития.
Модели динамики численности входят как важная часть в коммерческие модели развития агроценоза (P.M.Ives, A.B.Hearn, 1987). Такие модели объединяют как субмодели имитацию развития сельскохозяйственного растения и модели динамики численности наиболее важных вредителей. Помимо этих данных в компьютер регулярно вводится метерологическая информация. Модель дает информацию о прогнозируемом урожае, а также рекомендации о поливе полей и обработке их химическими средствами защиты растений (на основе порогов вредоносности). Нам неизвестны модели, которые давали бы рекомендации по сохранению полезных насекомых на поле или применению других биологических средств защиты растений.
Управление популяциями
Одна из основных практических задач современной экологии насекомых – разработка такой системы защиты растений, которая была бы основана на постоянном контроле и корректировке явлений, а не только на срочных, "пожарных" мероприятиях при выходе системы за установленные рамки. Эта система должна включать в себя как постоянное формирование желаемого агроценоза с помощью агротехнических мероприятий, например, подсева нектароносных растений, привлекающих многих паразитов, так и прямое регулирование численности прежде всего с помощью биометода. Это регулирование должно проводиться задолго до превышения численностью вида экономического порога вредоносности. Благодаря такому вмешательству уровень численности популяции останется на относительно низком уровне и не произойдет ее "ускользания" от пресса естественных врагов.
Для создания максимально устойчивой популяции желательно знать, насколько она способна сохранять свой средний уровень (буферность) и амплитуду колебаний около этого уровня (гомеостаз) (А. А. Шаров, 1985) по отношению к внешнему фактору. Отметим, что здесь речь идет о так называемых независимых по отношению к данной системе факторах. Всегда независимой будет, например, температура. В случае выпуска искусственно разведенных насекомых их численность тоже можно считать независимой величиной.
Применяющиеся в настоящее время интегрированные системы защиты растений – важный шаг вперед по сравнению с варварским использованием пестицидов, имевшим место ранее. При такой системе защиты растений решение об использовании того или иного метода борьбы принимается на основе сопоставления результатов учета численности с порогами экономической вредоносности. Естественно, что ожидаемая прибавка к урожаю в деньгах должна быть заведомо выше стоимости примененных средств защиты растений. В случае же применения химического метода величину прибыли следует также сопоставить с убытками, связанными с загрязнением среды, заболеваниями и гибелью домашних и диких животных, заболеваниями людей.
В ряде случаев вопрос о применении инсектицидов переходит из экономической в социальную сферу. Это очевидно в том случае, если инсектицид, накапливаясь, например, в воде, вызывает тяжелые заболевания и смерть людей, приводит к рождению дефективных детей или выкидышам. Совершенно очевидно, что руководители хозяйств, где необоснованно применяются такие инсектициды в больших масштабах, должны нести за это уголовную ответственность.
Интегрированная система защиты растений, как правило, предусматривает умеренное использование химических пестицидов или, что лучше, микробиометода, при превышении численности порогов вредоносности. Естественно, что на таком высоком уровне численности только такие радикальные способы защиты растений могут дать ощутимый эффект. Мы отмечали выше, что паразиты и хищники как фактор обладают высокой инерционностью, а незначительное снижение численности, вызванное ими, способно в ряде случаев даже продлить вспышку массового размножения вредителя.
Выпуск искусственно разведенных энтомофагов – это "тонкое" средство, способное предотвратить вспышку массового размножения вредителя в самом ее начале, когда численность вредителя еще низка, но уже начинает уходить от контроля естественных хищников и паразитов. В этой ситуации не требуется "наводнения" поля или леса разведенными на биофабрике паразитами и хищниками, поэтому стоимость регулирования не может быть высокой. Кроме того, потребуются усилия для максимального сохранения природных энтомофагов. Такая экологическая защита растений будет основана не на экономических порогах вредоносности, а иметь своей основной стратегической задачей сохранение естественной устойчивости агроценоза. При этом главные затраты будут направлены на мониторинг и обработку данных. Химические же средства защиты растений как нарушающие естественное равновесие агроценоза должны быть исключены.
Таким образом, в ближайшем будущем экологические системы защиты растений придут на смену интегрированным. Эти системы будут истинным управлением биоценозом.
ГЛАВА 4. НАСЕКОМЫЕ В ЭКОСИСТЕМАХ
Все живые организмы находятся в постоянном взаимодействии друг с другом и с неживой (косной) средой. На каждом участке пространства складывается более или менее устойчивый комплекс организмов и стабильный круговорот веществ между живой и неживой средой. Этот круговорот основан на притоке энергии извне (солнечной энергии). Таким образом, организмы и окружающая их среда образуют экосистемы.
Возможно подразделение экосистем либо по занимаемому ими пространству, либо по структуре связей между организмами. В принципе, эти подразделения переходят одно в другое.
В зависимости от величины охватываемого пространства экосистемы могут быть различных рангов, причем экосистема большего ранга представляет собой комплекс экосистем более низших рангов.
Высший ранг экосистемы – биосфера. Далее по рангу следуют экосистемы ландшафтных зон (Чернов, 1975: тундра, тайга, степь), материков и островов. Наиболее часто рассматривается экосистема одного из низших рангов, складывающаяся на определенной обычно относительно небольшой территории с более или менее однородными почвенными и климатическими условиями и обладающая определенным растительным сообществом (например, осоковое болото, сосновый лес, яблоневый сад, пшеничное поле). Такая территория носит название биотоп, все что на ней обитает и растет называют биоценозом, а в целом такая экосистема обычно именуется биогеоценозом (строго говоря, понятие "экосистема" несколько шире, так как включает в себя не только внутренние, но и внешние взаимодействия обитателей биотопа).
Население какого–либо вида животных в пределах биотопа рассматривается как популяция.
Биоценоз – это сложная система, состоящая из продуцентов (растений) или других источников органического вещества (например, кучки навоза), консументов (фитофагов и хищников) и редуцентов. Поэтому недопустимы такие термины как "энтомоценоз" или "фитоценоз" (К.В. Арнольди, Л.В. Арнольди, 1963). Можно, однако, говорить о комплексах насекомых, входящих в тот или иной биоценоз, или о населении.
Дата добавления: 2015-05-16; просмотров: 1224;