Фазовый портрет динамики численности 4 страница
Конструктивные сукцессии возникают после гибели по каким-то причинам существенной части биоценоза, прежде всего его растительного компонента, и длятся десятилетиями, а иногда столетиями и больше. Деструктивные же сукцессии требуют для полного цикла от 10–20 лет при разрушении древесины до нескольких суток, когда идет распад экскрементов. Смена стадий сукцессии протекает наиболее быстро на первых ее этапах.
Конструктивные сукцессии
Эти сукцессии начинаются с заселения мест, освободившихся после лесного пожара, наводнения, вырубки леса, лавин и геологических сбросов и по другим причинам.
Первые сообщества, появившиеся на таких местах, называют пионерными. Они состоят в основном из растений и животных г–стратегов. Различные флористические, а вместе с ними и фаунистические комплексы сменяют друг друга как бы в эстафете, пока сообщество не доходит до состояния климакса – окончательной своей стадии. На фоне таких сукцессии возникают циклические сукцессии более мелкого порядка. Так, известно, что для средней полосы нашей умеренной зоны климаксным сообществом могут быть дубовые леса. Однако после вырубки леса первыми на этом месте появляются мелколиственные древесные породы, обычно береза и осина. Под покровом такого леса вырастает ельник, постепенно вытесняющий лиственные породы. Но ели неустойчивы к сильным ветрам и на месте ветровала опять вырастает березняк. Долго живущие дубы на фоне этой смены растительности лишь постепенно завоевывают древесный полог. Естественно, что со сменой древесных пород меняются все консорции и с ними большая часть населения насекомых. Этот процесс захватывает столетия.
Насекомые–фитофаги играют активную роль в сукцессиях, значительно их ускоряя. Это происходит потому, что насекомые в первую очередь устраняют ослабленные растения. Переработка насекомыми древесных и других растительных тканей приводит к быстрому возвращению органических веществ в почву, что повышает устойчивость всего биоценоза. Насекомые способствуют также быстрой замене неустойчивых лесных и полевых монокультур, созданных человеком, на более сложные и устойчивые естественные биоценозы.
В результате постоянной деятельности человека во многих случаях климакс не достигается, но возникает другое устойчивое состояние – дисклимакс. Например, чрезмерный выпас скота может превратить степь в пустынное сообщество с соответствующей, хотя и несколько измененной и обедненной флорой и фауной. Дисклимаксными можно считать и относительно устойчивые сообщества, возникающие на пустырях и характеризующиеся рудеральной растительностью. Иногда к этой категории относят и агроценозы, которые неоднократно за год подвергаются воздействию человека. Естественные фитофаги и сорняки быстро могут превратить их в типичные для данного климата пионерные сообщества с преобладанием растений и животных r–стратегов.
Деструктивные сукцессии
Все деструктивные сукцессии представляют собой разложение органического субстрата до почти полной его минерализации. Эти сукцессии легко наблюдать на примере постепенного изменения опада растительности – подстилки. Так как опад поступает в подстилку более или менее регулярно, то в подстилке возникает слоистость, содержащая субстрат на разных стадиях разложения и, соответственно, с разным набором бактериальной флоры и фауны. В целом для подстилки характерно большое экологическое разнообразие (Н.М.Чернова, 1977). Здесь обитают сотни видов нематод и членистоногих. В число последних входят клещи, ногохвостки, личинки различных двукрылых, жуки. В подстилке низка степень доминирования массовых форм и имеется большое количество видов с устойчиво низкой численностью.
Наблюдения за сукцессиями в составе фауны подстилки осложняются миграциями членистоногих, связанными со степенью увлажнения подстилки (погодой) и сезоном. В течение сукцессии четко прослеживается все же постепенная смена поверхностно живущих форм членистоногих подстилочно–почвенными, а затем почвенными формами.
Время, необходимое для минерализации и гумификации хвойного и лиственного опада в умеренном климате, равно примерно 9–12 годам. Членистоногие играют существенную роль в этом процессе, механически измельчая субстрат и пропуская его сквозь кишечник, где он вместе с бактериальной и грибной флорой обрабатывается пищеварительными ферментами.
Иные комплексы видов насекомых принимают участие в разложении древесины, например упавших стволов деревьев или пней. Можно различить 4 стадии этого процесса (Б.М.Мамаев, 1960). Первая стадия имеет место, когда отмершая древесина остается еще твердой, хотя в ней уже поселяются различные деревоокрашивающие грибы. На этой стадии в древесине живут личинки разных усачей, а слой коры интенсивно потребляется жуками–короедами и личинками усачей–рагиев. На второй стадии древесина приобретает бурый цвет и легко раскалывается на пластинки. Дерево продолжают разрушать усачи, а позже к ним присоединяются личинки жуков–рогачей. На третьей стадии древесина становится очень ломкой и имеет красный цвет. Основное население такой древесины – муравьи. Упавшие стволы на этой стадии разложения обычно покрыты мхом. Наконец, древесина превращается в темно–коричневую труху, заселенную дождевыми червями, многоножками, личинками комаров Bibionidae и другими насекомыми. Отметим, что при разложении древесины отдельные комплексы фауны могут и выпадать, хотя итог разложения древесины всегда один и тот же.
Значительно быстрее протекает сукцессия на субстратах, богатых азотом. В целом на первых стадиях разложения любых таких субстратов раньше других появляются яйца и личинки мух (особенно Muscidae, Calliphoridae, Sarcophagidae), некоторые жуки и форезирующие членистоногие, в основном клещи, заносимые этими насекомыми. Постепенно они замещаются почвенно–подстилочными видами и, наконец, почвенными. Различия в видовом составе, связанные с типом субстрата, очень заметны на первых этапах разложения, а потом постепенно сглаживаются. Правда, при низкой влажности на сухих остатках трупов очень долго сохраняются жуки–кожееды и моли.
Н.М.Чернова описывает следующие изменения во времени фауны конского навоза. Навоз, пролежавший не более суток, содержит много мелких нематод, яйца мух–дрозофил, некоторых форезирующих клещей. На 3–й день в навозе помимо той же фауны, начинают попадаться личинки жуков–стафилинид. На десятый день могут быть обнаружены личинки водолюбов (Cercyon) и взрослые жуки тех же видов, а также жуки–карапузики. В двенадцатидневном навозе обильны ногохвостки (Hypogastrura) а также мелкие жуки семейства Ptiliidae.
На богатых органических субстратах возникает жесткая конкуренция между его потребителями. Так, навоз может очень быстро утилизироваться жуками. В южных районах скарабеи и некоторые другие навозники делают из навоза шары, которые катят с открытого места в специально делаемые укрытия, позволяющие избавиться от конкурентов. Жуки–могильщики Necrophorus с той же целью зарывают трупы мелких животных в почву.
Очень сложная и напряженная конкуренция между насекомыми, грибами и нематодами требует от первых заселителей большой скорости развития, что облегчается разогревом субстрата до 30–40° С. Так, комнатная муха в свином навозе способна развиться от свежеотложенного яйца до куколки за 5–6 суток. Большое значение имеет групповое внекишечное пищеварение, выделение ряда веществ, убивающих конкурентов или тормозящих их рост, и также изменения рН субстрата. Естественно, что большая концентрация насекомых в быстро разлагающихся субстратах привлекает многих хищников и паразитов. Даже среди Muscidae есть виды, личинки которых поедают личинок мух других видов. Первая стадия разложения субстрата, богатого белками, длится не более нескольких дней. Далее следуют другие менее напряженные в отношении конкуренции стадии, где практически полностью сменяются все группировки насекомых.
6. АНТРОПОГЕННЫЕ ЭКОСИСТЕМЫ
Хозяйственная деятельность человека приводит к резким изменениям всех компонентов биоценозов, в том числе и насекомых (М.В.Козлов, 1990). При этом многие виды вымирают или становятся редкими. Существенно сокращаются и дробятся на части ареалы многих видов. Некоторые же виды получают преимущества и становятся массовыми. Кроме того, в ряде случаев фауна обогащается новыми видами, бессознательно или сознательно завезенными человеком. Степень и характер изменений биоценозов определяются родом и интенсивностью его хозяйственного использования (С.И. Медведев, 1959).
Так, умеренный выпас скота не наносит существенного вреда луговому и степному биоценозу, поскольку копытные – естественный компонент многих биоценозов, повышающий их устойчивость. Однако слишком интенсивный выпас приводит к обеднению флоры и, соответственно, фауны насекомых при общем сдвиге всех компонентов в сторону большей ксерофильности. Выпас скота – это не только изъятие части зеленой массы, но и нарушение почвенного покрова, особенно его поверхностной структуры, появление многочисленных кровососущих насекомых и обитателей навоза. Интересно, что в некоторых случаях сохранению прежней фауны насекомых существенно способствуют колючие кустарники и полукустарники, под прикрытием которых остаются нетронутыми многие другие растения (B.C. Мурзин, 1981).
Снижение видового разнообразия происходит и при кошении трав, которое приводит к выпадению из состава флоры ряда однолетних и двулетних растений, которые лишаются возможности давать семена. На покосе погибает масса личинок насекомых, особенно обитающих внутри стеблей. Интересно, что только ежегодное скашивание трав резко обедняет состав фауны. При нерегулярном сенокошении комплекс насекомых заметно обогащается, так как, оно, по–видимому, в какой–то мере заменяет отсутствие или недостаток диких копытных или же умеренный выпас скота. В абсолютно заповедной степи, где запрещены и выпас, и сенокошение, видовой состав насекомых заметно обеднен и они представлены в основном прямокрылыми и некоторыми клопами.
Рекреационная нагрузка на биоценозы, сопровождающаяся вытаптыванием и замусориванием местности, влияет на фауну подобно интенсивному выпасу скота, существенно ее обедняя.
Своеобразная ситуация возникает на обочинах дорог. С одной стороны, частые повреждения и замусоривание приводят к расцвету рудеральных (мусорных) растений (лопух, крапива, полынь, пустырник и т.д.) с их своеобразной фауной, с другой – именно около дорог иногда сохраняются остатки прежней целинной флоры и фауны (С.И. Медведев, 1959). С дальнейшей урбанизацией местности эти остатки, как правило, исчезают.
Вырубка леса, а тем более распашка целины приводят к исчезновению большинства элементов прежнего биоценоза. На полях и в садах возникают искусственные сообщества, создаваемые и постоянно поддерживаемые человеком, – агробиоценозы, о которых речь пойдет ниже. Урбанизация местности приводит к полному уничтожению прежних биоценозов.
Своеобразные экосистемы, состоящие практически из одного вида, создаются человеком при массовом разведении насекомых. Такие лабораторные экосистемы иногда называют техноценозами (А.З. Злотин, 1989).
Агробиоценозы
Агробиоценоз (агроценоз) – пример вторичного или дисклимаксного биоценоза. Он отличается тем, что в нем искусственно поддерживается резкое доминирование (обычно фактически монокультура) какого–либо одного сельскохозяйственного растения. Ежегодно часть биомассы, создаваемой агроценозом, изымается в качестве урожая, при этом агроценоз теряет как органические, так и минеральные вещества.
Такое искусственное образование, как агроценоз, может сохраняться только при постоянном и ежегодном возобновлении его человеком. Территория любого брошенного агроценоза быстро заселяется сорной растительностью, культурные же растения с первых лет оказываются третьестепенным компонентом флоры, а потом исчезают.
Однако в течение периода вегетации на поле формируется сложный и иногда хорошо сбалансированный комплекс насекомых, насчитывающий сотни видов. В этот комплекс входят фитофаги, повреждающие культурные растения, фитофаги, съедающие сорняки, и многочисленные энтомофаги. Поэтому хорошо укоренившееся и весьма распространенное представление, что "агроценоз не обладает свойством регуляции" (Л.В.Арнольди, К.В. Арнольди. 1963), уже нанесло и продолжает наносить нашему и мировому хозяйству колоссальный вред. Вместо контроля за агроценозом и попыток повысить его устойчивость, пассивно ожидается наступление массового размножения вредителя, а затем применяются радикальные меры. Такая защита растений тоже сохраняет урожай, но при этом расходуется много средств и обычно загрязняются ядами пищевые продукты, вода и все окружающие биоценозы.
Естественная устойчивость агроценоза может быть значительно повышена за счет граничащих с ним биоценозов, часто являющихся постоянным источником хищников и паразитов (О.Д.Ниязов, 1992; А.Ш.Хамраев, 1992). Если эти биоценозы находятся не на пионерных стадиях сукцессии, на них должны отсутствовать фитофаги – r–стратеги, многие из которых являются вредителями сельскохозяйственных растений. Следовательно, чем устойчивее биоценоз, прилегающий к полю с сельскохозяйственной культурой, чем менее пионерным он является, тем больше пользы он может принести соседним сельскохозяйственным угодьям.
К сожалению, в сельском хозяйстве обычно применяется обратная практика. С целью устранения сорняков и вредителей часто перепахивают обочины дорог, а в некоторых случаях их даже выжигают. Естественно, что на таком оголенном участке кроме растений и животных r–стратегов никто поселиться не может, и эти участки являются поставщиками сорняков и вредителей полю. Более правильным было бы или совсем не трогать эти участки, или же ускорять на них сукцессию с помощью семян растущих в устойчивых естественных сообществах трав (посев мелко нарезанного сена).
Значительно повышает устойчивость агроценоза также включение в посев каких–либо дополнительных культур (совместный посев – В.Г.Коваленков, Н.М.Тюрина, 1993) или посев отдельными полосами разных растений вместо больших полей монокультур. Введение в посев цветущих растений, например, их посев по обочинам вокруг поля, обеспечивает дополнительное питание паразитам и хищникам и положительно сказывается на урожае. Усложнение агроценоза не только увеличивает численность энтомофагов на поле, но и улучшает микроклимат посевов.
Формирование комплекса вредителей на поле – довольно сложный процесс. При распашке целинной земли ее обитатели в основном погибают или мигрируют. Причины их гибели следующие: изменения структуры почвы, ее температурного и влажностного режима, исчезновение кормовых растений – центров консорций, а также воздействие агротехнических приемов. Ряд насекомых все же приспосабливается к жизни на посевах. Среди них виды, размножающиеся вне пределов поля, но охотно питающиеся культурными растениями (многие саранчовые), насекомые, мигрирующие на зиму на опушки леса, на обочины (например, клоп–черепашка), обитатели взрыхленных грызунами почв в природных биотопах (например, жуки Anisoplia austriaca Host., Pentodon idiota Hbst., озимая совка Agrotis segetum Schiff.). В эту же группу можно включить и обитателей растений таких взрыхленных почв (многие клопы и листоблошки). На полях оказываются также насекомые, предпочитающие изреженный травостой (щелкуны родов Melanotus, Agriotes, чернотелки Opatrum sabulosum L.). Весьма распространены и насекомые, зимующие в многолетних или озимых растениях или в растительных остатках (двукрылые Oscinosoma, Chlorops, Mageti–ola).
Таким образом, фауна поля формируется в основном за счет местных видов. Так, при вспашке целинных земель в Казахстане в 50–е годы доминантными видами на пшенице оказались пшеничный трипе Нарlothrips tritici Kurd., серая зерновая совка Hadena sordida Bkh., сибирский остроголовый клоп Aelia sibirica Reutt., хлебная полосатая блоха Phyllotreta vittula Reutt.). Все эти олигофаги обитали до распашки на степных злаках (Т.Г. Григорьева, 1970). Резкое увеличение пищевых ресурсов и более высокая пищевая ценность культурной пшеницы, по сравнению с зернами диких злаков, явились причиной небывало массового размножения серой зерновой совки в первые годы освоения целины. По–видимому, в это время произошло "ускользание" этого вида из под пресса паразитов и хищников. Однако через 5–6 лет возделывания пшеницы на целинных землях возник новый уравновешенный комплекс насекомых. При этом значительно увеличилось число видов насекомых на полях, снизилась их общая численность и возросла роль энтомофагов.
В случае заселения целинных земель, лежащих поблизости от окультуренных (хлопчатник – Ю.И. Алексеев и др., 1976), происходит постепенное заселение новых посевов без вспышек массового размножения. В результате также формируется устойчивый комплекс насекомых.
Итак, на культурных растениях поселяются местные олигофаги и полифаги. Если же растение чуждо местной флоре и поблизости нет родственных ему видов, его фитофагами становятся только многоядные насекомые. Примерно так происходит заселение некоторых лесопосадок. Заметим, что переход насекомых на новые для них растения осуществляется во многих случаях очень постепенно, что, по–видимому, связано с выработкой новых пищевых рас (М.С. Гиляров, 195S).
Агротехнические мероприятия существенно сказываются на фауне насекомых. Так, снегозадержание явно улучшает перезимовку куколок озимой совки. При орошении исчезают степные ксерофилы но увеличивается численность шведской мухи, хлебных блох, злаковых цикадок. В результате севооборота заметно снижается численность ряда видов, но видовое разнообразие возрастает, так как каждая сельскохозяйственная культура оставляет свой след в энтомофауне (С.И. Медведев, 1959). Смена растений приводит к тому, что среди почвенных личинок, развивающихся более одного года, выживают практически только полифаги: проволочники, личинки чернотелок и пыльцеедов (М.С. Гиляров, 1945). Монофаги же, такие как личинки жуков–усачей Dorcadion, развивающиеся два года, совсем не могут перенести смену культур. Наоборот, быстро развивающиеся и хорошо мигрирующие насекомые, к числу которых относится много вредителей, практически не страдают от севооборота.
Остановимся вкратце на понятии вредоносности. Весьма распространена наивная точка зрения, что любое насекомое, поедающее культурное растение, уменьшает урожай и наносит таким образом вред. Однако между растением и насекомым исторически установились сбалансированные взаимосвязи, позволяющие сохранять эти растения как источники корма (В. И. Танский, 1988). Пока уровень повреждения относительно низок, вся популяция растений и отдельные их особи способны компенсировать ущерб. Иногда повреждения стимулируют растение, приводя к его более мощному развитию. Вообще практически всегда растения формируют больше вегетативных и генеративных органов, чем это необходимо для оптимальной продуктивности. Так, у хлопчатника наблюдается естественный опад до 80% цветков и завязей. При искусственном повреждении цветов и завязей этот естественный опад резко уменьшается и общий урожай хлопчатника увеличивается.
Даже в том случае, если насекомое приводит растение к гибели, что типично для вредителей всходов и семян в почве, изреживание посевов компенсируется более мощным развитием сохранившихся растений. Например, при гибели 25% всходов свеклы от проволочников, общий урожай не снижается (С.Г.Бобинская и др., 1965). Сравнивая урожай растений, неповрежденных и поврежденных вредителем, можно определить коэффициент вредоносности:
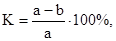
где а – масса урожая неповрежденных растений и b – масса урожая того же количества поврежденных растений.
Если слабые повреждения могут привести к увеличению урожая, то с дальнейшим ростом плотности популяции вредителя масштаб повреждений будет увеличиваться и начиная с определенного уровня плотности приведет к потере урожая (порог вредоносности). Но любые защитные мероприятия связаны с расходами, которые зависят не только от стоимости препаратов или агентов биологической защиты растений, включая их применение на поле, но должны отражать и ущерб, наносимый людям (например, повышение заболеваемости и оплату бюллетеней), загрязнение окружающих угодий, в том числе водоемов, в которых рыба погибает от химических препаратов, ухудшение структуры почвы, что приводит к дальнейшему снижению урожайности, а в ряде случаев, и снижение закупочных цен на продукт, содержащий остатки инсектицида.
Поэтому в нашей стране считается целесообразным применение химических средств только в том случае, если затраты на применение инсектицида будут не менее, чем троекратно, окупаться прибавкой урожая (В.И.Танский, 1988). Такой уровень потерь достигается если численность вредителя приближается к так называемому экономическому порогу вредоносности. Соотношение порога вредоносности и экономического порога вредоносности показано на рис. 42. Экономические пороги вредоносности определены для большинства основных сельскохозяйственных культур и многих вредных насекомых. Разработка этих порогов – большое достижение в защите растений, позволяющее уменьшить как затраты, так и загрязнение среды. Эти пороги могут меняться в зависимости от конкретной ситуации, прежде всего от фазы развития растения. Примеры подобных порогов для зерновых культур и клопа – вредной черепашки приведены в табл. 4.
Однако возможности работы в этом направлении еще далеко не исчерпаны по следующим причинам.
Во-первых, большое количество энтомофагов на поле делает обработку пестицидом бесполезной, а, точнее, вредной даже когда численность фитофага существенно превышает экономический порог.
Таблица 4
Экономические пороги вредности клопа вредной черепашки на пшенице (В.И.Танский, 1988)
| Стадия насекомого | Фаза развития растений | Экономический порог вредности |
| Перезимовавшие клопы | отрастание – кущение (озимая пшеница) | 1-2 клопа/м 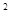
|
| кущение (яровая пшеница) | твердая пшеница; 0,3-1,1 клопа/м
мягкая пшеница: 0,5-1,5 клопа/м
| |
| Личинки | цветение – начало налива зерна | 5-10 личинок/м
|
| молочная спелость | сильная и ценная пшеница:
1-2 личинка/м
рядовая пшеница:
5-6 личинок/м
|
Следовательно, вместе с численностью фитофага необходимо определять и численность его основных энтомофагов (Н.В.Бондаренко, 1986).
Во-вторых, высокая плотность популяции фитофага часто приводит к развитию эпизоотии, снижающих его численность не хуже любого инсектицида. Следовательно, необходимо определять также и процент заболевших насекомых.
В-третьих, экономические пороги определены для каких-то "усредненных" растений данной сельскохозяйственной культуры и для усредненных метеоусловий. Очевидно, что способность растения самостоятельно компенсировать ущерб должна зависеть от сорта, сроков посева, геохимического состава почвы, внесения удобрении, поливов, а также погоды.

Рис.42. Зависимость урожая возделываемого растения от плотности популяции вредителя. 1 – порог вредоносности, 2 – экономический порог вредоносности (с учетом расходов на проведение защитных мероприятий и ущерба окружающей среде, включая возможное загрязнение продукта питания) (по В.И. Танскому, 1988)
Кроме того, эти условия могут влиять на популяцию насекомых или непосредственно, или косвенно – через изменения физиологического состояния растений. Поэтому даже для одной культуры и одного и того же вредителя в зависимости от условий кривые, показанные на рис.42, сохраняя ту же форму, пойдут совершенно по-разному (Л.Е.Славгородская–Курпиева, 1990)
Таким образом, определение экономических порогов вредоносности – это начало пути, который на базе объединения усилий энтомологов, физиологов растений, почвоведов приводит к созданию компьютерных моделей. Целью таких моделей является оптимизация сельскохозяйственного процесса для получения максимального урожая при наименьших затратах и минимальном загрязнении среды и продукта. Имея такую программу, в компьютер вводят информацию о состоянии растений и погоде. Компьютер же на экране монитора дает прогноз о будущем урожае и рекомендацию по агротехническим и защитным мероприятиям, которые желательно провести на данном поле и в данный момент времени.
Мы полагаем, что современная интегрированная защита растений, основанная на экономических порогах вредоносности, в ближайшие годы будет заменена экологической системой защиты растений. Эта система будет иметь профилактический характер и базироваться на динамичных порогах ускользания. Компьютерные модели экологической зашиты растений смогут рекомендовать растениеводу, как сохранить естественную устойчивость агроценоза и минимальными усилиями не допустить развитие вспышки массового размножения вредителей.
Насекомые города
Городская застройка местности приводит к уничтожению природных биоценозов и к возникновению очень специфических новых. Эти городские биоценозы относительно бедны видами и довольно просты по своей структуре. Они всегда зависят от деятельности человека и автотрофы – растения, создающие органическое вещество, как правило, непосредственно в них не входят.
Если насекомое спонтанно обитает в поселениях человека против его воли, а также сосуществует с человеком и зависит от его деятельности, такое насекомое может быть названо синантропным. Степень синантропности вида можно определить по следующей формуле (P.Nuorteva, 1963):
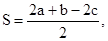
где а – доля особей данного вида среди сбора всех насекомых данной группы в поселении человека, b – то же в сельской местности в пределах агроценозов, с – то же в естественных биотопах. При значении индекса, близком к + 100, насекомое явно предпочитает городскую среду. Если индекс близок к нулю – поселения человека не влияют на популяции этого вида. При индексе – 50 и ниже вид явно избегает поселения человека.
Этот индекс синантропности различен для разных географических популяций одних и тех же видов. Например, на юге синантропные мухи и тараканы гораздо менее тесно связаны с жильем, чем на севере.
В городе создается конгломерат самых разнообразных условий. Естественно, что фауна отапливаемых помещений, подвалов, балконов, промышленных и складских строений, улиц и парков должна быть совершенно различной. Типичными для всех этих местообитаний является их мозаичность, расчлененность, что затрудняет миграции от одного местообитания к аналогичному другому (островной эффект – М.В. Козлов, 1990).
Но даже в пределах такого местообитания выявляются не связанные друг с другом группы видов. Так, в отапливаемых жилых домах можно встретить различных вредителей запасов (например, малого мучного хрущака, хлебного точильщика, зернового долгоносика, некоторых бабочек–огневок), вредителей материалов и изделий (например, домового усача, платяную моль, разнообразных кожеедов), паразитов человека (например, постельного клопа, комаров Culex pipiens molestus), паразитов домашних животных и синантропных грызунов (например, блох), обитателей плесени, а также водорослево–бактериальных нарастаний на "потеющих" в зимнее время трубах (некоторых ногохвосток, щетинохвостку – сахарную чешуйницу, насекомых, потребляющих остатки пищи и пищевые отходы (тараканов, некоторых дрозофил, некоторых муравьев), синантропных двукрылых, которые, однако, обычно размножаются за пределами помещения, фитофагов на комнатных растениях (прежде всего тлей и щитовок, некоторых клещей), а также паразитов этих фитофагов, детритофагов, живущих в почве цветочных горшков и под ними (ногохвостки, некоторые мелкие двукрылые).
Общим для всей фауны отапливаемых помещений является их теплолюбивость. Часто это адвентивные (занесенные) виды тропического происхождения. Все виды, обитающие в этих условиях, не имеют облигатной диапаузы, а иногда вообще не способны диапаузировать. Кроме того, все они, за редкими исключениями, легко переносят большую сухость воздуха.
Как правило, эти насекомые размножаются тут же в жилище. Уже упомянутым исключением являются синантропные мухи, а также размножающиеся в полузатопленных подвалах домов комары, нападающие на людей в помещениях даже в зимнее время. Постоянным источником многих кожеедов, а иногда и молей являются гнезда голубей, воробьев и других птиц, расположенные на чердаках и в других местах дома. Эти насекомые в массе развиваются, потребляя опавшие перья, трупы птиц, а иногда и остатки их пищи. Синантропные мухи и кожееды могут развиваться также на трупах крыс и мышей.
Значительно более подробный обзор фауны города, особенно жилых домов, дан М.В.Полежаевой (1984) на примере Москвы.
Относительно богата фауна парков, но и здесь заметно преобладают эвритопные, широко распространенные виды. Крупные насекомые в парках встречаются довольно редко и фауна представлена, в основном мелкими видами. Вообще чем ближе к центру города, тем меньше крупных насекомых там можно обнаружить. Среди городских насекомых нередко обнаруживаются виды, происходящие из более южных районов, что связано с более теплым климатом города, по сравнению с окружающей его местностью. В целом количество видов насекомых в центре города всегда меньше, чем на его окраине, а тем более, в сельской местности.
Дата добавления: 2015-05-16; просмотров: 929;