Фазовый портрет динамики численности 2 страница
Иногда используется также пространственное вертикальное подразделение биогеоценоза на ярусы. Так, можно говорить, например, о почвенном ярусе или о пологе леса. Здесь действительно образуются особые комплексы видов, тесно связанные друг с другом.
Микроучастки, отличающиеся особыми условиями от всего биотопа (поверхность листа или ствола, пень, труп животного, плодовое тело гриба) называют микробиотопами. Соответственно, весь комплекс растений, грибов, животных, заселяющих микробиотоп, может быть назван микробиоценозом.
Очень близко к предыдущему разделение насекомых на комплексы по их местообитанию. Так, в монографии В.Г.Каплина (1981) рассмотрены, например, комплексы насекомых и других членистоногих в древесине деревьев, в галлах, цветках, плодах и семенах.
Среди экосистем, рассматриваемых в плане взаимосвязей между живыми объектами, выделяют консорции. В пределах консорции все организмы четко объединены между собой трофическими связями. Обгоняющий центр консорции обычно – свободно живущее автотрофное растение, являющееся источником органического вещества для всех прочих организмов, от фитофагов и до сверхпаразитов. В консорцию входят также организмы, использующее это растение в качестве укрытия. Иногда в качестве центра консорции принимают животное (В.Н.Беклемишев, 1970) со всем специфическим комплексом членистоногих и других организмов. Консорции обычно складываются из более тесных группировок, возникающих, например, на грибе–трутовике, растущем на стволе дерева, в галлах, на соке, вытекающем из поврежденного ствола, и, наконец, на различных частях растения. Можно также говорить о консорциях на отмерших гниющих деревьях, а также на трупах животных и их экскрементах, на плодовых телах грибов. Такие группировки одновременно имеют четко выраженные пространственные границы и совпадают с микробиоценозами.
В экологии используется также представление о жизненной системе популяции, центром которого является популяция изучаемого вида. Жизненная система включает в себя все виды организмов, с которыми изучаемый вид вступает в тот или иной контакт, а также абиотические факторы, влияющие на жизнь популяции.
Иногда рассматриваются также так называемые коадаптивные комплексы (Г.М.Длусский, 1981), под которыми понимают совокупность популяций разных видов, входящих в один биогеоценоз и связанных конкурентными или мутуалистическими отношениями. Г.М.Длусский описывает коадаптивные комплексы на примере муравьев пустыни, рассматривая их как наборы определенных жизненных форм.
В настоящей главе мы уделим основное внимание биогеоценозам, а также консорциям и роли в них насекомых. Насекомые могут выступать как фитофаги, зоофаги и сапрофаги. При изучении роли насекомых в любом биогеоценозе начинают с определения их видового состава, а затем устанавливают положение данных насекомых в системе биогеоценоза и их взаимосвязи с другими видами. Возможны также три количественные характеристики данного вида: численность (плотность популяции), биомасса и поток энергии, проходящий через особей данного вида.
1. ИЗУЧЕНИЕ ВИДОВОГО СОСТАВА
Следует различать фаунистические и демэкологические аспекты исследований. Фаунистика изучает географическое распределение видов или других систематических единиц, а демэкология – географическое распределение сообществ (экосистем). В первом случае мы вправе говорить о фауне, а во втором – о животном населении. Видовой состав – важнейшая характеристика животного населения (Ю.И. Чернов, 1984). Исследование любой экосистемы начинается с установления систематической принадлежности входящих в нее животных, т.е. составления списка видов или более крупных систематических единиц. Очевидно, что наиболее информативный материал можно получить при установлении точного видового состава. Однако на практике из-за технических причин и большой трудоемкости определения отдельных групп иногда ограничиваются определением до семейства или даже отряда.
Хотя экология близких видов, обитающих вместе в одном и том же биотопе, всегда различна (правило Г.Ф.Гаузе), по отношению к какому-то отдельно взятому фактору их реакция может быть однозначной. Если реакция на фактор проявляется у группы не различаемых нами видов, принадлежащих к одному более высокому таксону (суммарный учет), это означает, что либо все виды проявляют одинаковую реакцию, либо ее проявляет только массовый (или массовые) вид. Если же реакция при анализе суммарного обследования отсутствует, то ее или вообще нет у представителей этой группы видов, или она проявляется в разных направлениях у нескольких массовых видов и, таким образом, при суммарном анализе нивелируется.
Видовой состав может быть изучен в пределах экосистемы любого ранга. Однако здесь всегда следует учитывать, что фауна каждого региона помимо чисто экологических причин, определяется историей ее генезиса. Для фауны региона это может быть история на протяжении геологических периодов. Но даже в биотопах при их смене тоже длительно сохраняется отпечаток прежней фауны. Так, при организации Ботанического сада МГУ на месте бывших колхозных полей полевые виды насекомых сохранялись там более 10 лет, несмотря на смыкание древесной и кустарниковой растительности.
Никогда нет гарантии, что выявлен полный видовой состав в пределах данного таксона на данной территории, не только в силу неизбежных пропусков, но также из-за потока постоянно прибывающих мигрантов. Например, в Казахстане был отмечен занос мигрирующими птицами мух–кровососок из очень отдаленных тропических районов (Т.Н.Досжанов, 1982). Конечно, чем больше применено разных методов сбора и чем дольше проводилось исследование, тем более вероятно, что видовой состав практически исчерпан. В случае сборов насекомых с помощью проб четко выявляется быстрое исчерпание видового состава с выходом графика зависимости числа видов от количества проб к асимптоте уже через 15–20 проб.
Количество видов в биоценозах обычно возрастает при продвижении с севера на юг. Однако это правило далеко не абсолютно. Количество видов насекомых, по–видимому, определяется как суровостью условий, так и степенью разнообразия растительного покрова. Так, в пустынях количество видов может быть меньше, чем в более северных и увлажненных районах с богатой и разнообразной флорой. Кроме того, количество видов зависит от стадии сукцессии, т.е. этапа развития биоценоза. Оно, как правило, мало в нарушенных по каким–либо причинам биоценозах, находящихся на ранних стадиях своего развития.
При обследовании нескольких экосистем одного ранга, как и при изучении фауны нескольких географических регионов, возникает необходимость сравнить их друг с другом. Это делается с помощью вычисления следующих коэффициентов:
коэффициента Жаккара 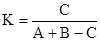 и
и
коэффициента Серенсена 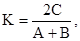
где А – число видов данной систематической категории в одной группировке, В – то же в другой, а С – число видов, общих для обеих группировок. В исследованиях часто сравнивают группировки попарно с помощью обоих коэффициентов. Коэффициент Жаккара, в принципе, более строг и пропорционален, а коэффициент Серенсена имеет большую "разрешающую силу" при низких величинах сходства. На практике эти коэффициенты попарного сходства представляют в виде таблицы спортивных соревнований, где попарно встречаются разные команды, причем верхнюю правую половину заполняют индексом Жаккара, а нижнюю левую – индексом Серенсена.
Список видов далеко не является исчерпывающей характеристикой экосистемы. В пределах двух биотопов могут существовать одни и те же виды, но соотношение их численностей может быть различным. О численности как важнейшей характеристике популяции мы уже говорили выше.
Напомним, что самый простой способ ориентировочно оценить численность, – это определить встречаемость, т.е. процент проб в которых встречен данный вид, независимо от количества особей в пробе. Отметим, что наиболее точную оценку этот показатель дает при случайном размещении на местности особей или их небольших, по сравнению с пробой, скоплений. При агрегированном размещении точность этого показателя снижается.
Процент проб с данным видом имеет смысл определять лишь в том случае, когда все пробы примерно равны по охватываемой площади. На практике же часто приходится иметь дело со многими сборами разных авторов, обычно неравноценными как по объему, так и по стилю сбора материала. Однако даже из таких данных можно вынести общее представление о редкости тех или иных видов, определяя количество особей интересующих нас видов во всех сборах (Д.В. Панфилов, 1976).
Предположим, что имеется всего 20 неравноценных сборов с одной территории. Один вид, который мы считаем редким, представлен 50 экземплярами в одном сборе. Другой же вид насчитывает в трех сборах по 5 экземпляров, а в остальных – по 10. Индекс встречаемости равен:
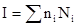 ,
,
где n 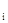 – число особей в сборе, а N – число сборов, где имеется n экземпляров.
– число особей в сборе, а N – число сборов, где имеется n экземпляров.
Тогда для первого вида индекс равен I = 50 х 1 = 50, а для второго:
I = 5 х 3 + 10 х 17=185
Подчеркнем, однако, что абсолютно недопустимо сравнивать фауны каких–либо географических районов по однократным сборам, сколь бы они не были обильны. В самом деле, соотношение численностей видов меняется от сезона к сезону, при этом фенологические события могут изменяться по датам. Кроме того, соотношение численностей разных видов из года в год существенно меняется. Только многократные сборы, проведенные в разные сезоны и в разные годы, дают более или менее точное представление о фауне данного географического района.
В отличие от встречаемости, плотность популяции, т.е. количество особей данного вида на единицу площади, является более устойчивым экологическим показателем, в принципе, не зависящим от метода учета. Очевидно, что биоценозы отличаются друг от друга не только по видовому составу, но и по плотности популяций входящих в них видов (обилию). Поэтому более информативное представление о различиях биоценозов можно получить, используя формулу Р. Л. Наумова (1964):
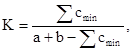
где а – суммарная плотность популяций всех видов, обнаруженных в первом биоценозе, b – то же для второго биоценоза, а с – минимальный показатель плотности популяции вида, встречающегося в обоих биоценозах.
Число массовых видов в биоценозе никогда не бывает большим. В устойчивых биоценозах наибольшее количество видов обладает умеренной численностью. Очень же редких видов, представленных в обширных сборах одной–двумя особями, тоже сравнительно немного. Поэтому если мы на оси абсцисс отложим количество пойманных особей, выраженное в логарифмах, а на оси ординат – количество видов, представленное данным числом особей в сборах, то получим колоколовидную кривую, сходную с классической кривой нормального распределения (логарифмически–нормальное, или лог-нормальное распределение). В случае загрязнения биотопа или деградации сообщества по другим причинам уменьшается как численность всех или почти всех видов, так и общее число видов и кривая сдвигается на графике влево, т.е. оказывается больше всего видов, представленных малым числом особей (распределение, близкое к геометрическому или же логарифмическому ряду).
В последнее время особое значение придают индексам видового (экологического) разнообразия (Э.Мэгарран, 1992). Эти показатели складываются из двух компонентов. Первый из них – это видовое богатство. Его можно определить как отношение общего числа видов к общему числу особей в пробах. На практике часто используют другой индекс видового богатства, предложенный Р.Маргалефом:
D = 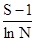
где S – число выявленных видов, а N – общее число особей всех видов.
Второй компонент – это выравненность распределения особей между видами. Ю.Одум (1975) приводит следующий схематический пример: "... две системы, каждая из которых состоит из 10 видов и 100 особей, могут иметь совершенно одинаковые отношения (видового богатства, которое и в том, и в другом случае равно 9/100 –В.Ч.)..., но совершенно разные показатели выравненности в зависимости от распределения 100 особей между 10 видами. Так, в одном случае это распределение может иметь вид: 91–1–1–1–1–1–1–1–1–1 (минимальная выравненность), в другом случае – по 10 особей на вид (максимальная выравненность). Некоторые индексы видового разнообразия учитывают оба эти параметра. Здесь мы приводим формулу только одного, наиболее часто применяемого индекса Шеннона (Э.Мэгарран,1992):
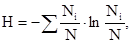
где Н – видовое разнообразие в битах, Ni – число особей каждого вида во всех пробах, N – общая численность особей всех видов во всех пробах. Соотношение численности данного вида и суммарной численности всех видов (N / N) называют коэффициентом доминантности данного вида. Индекс Шеннона обычно меняется в пределах от 1,5 до 3,5.
Как мы уже отмечали выше, со снижением видового разнообразия резко меняется видовая структура сообщества. При этом часто резко повышается обилие отдельных немногочисленных видов (биоценотический закон Тинемана – A.Thieneman, 1925). Легко убедиться что там, где максимально флористическое богатство, много видов с малой плотностью популяции и совсем отсутствуют многочисленные доминирующие виды. Для антропогенных же ландшафтов типично обеднение видового состава и проявление отдельных резко доминирующих видов.
Индексы видового разнообразия применяются для мониторинга окружающей среды, а также при выборе участков для создания заповедников. При этом предполагается, что высокие показатели разнообразия свидетельствуют о наименьшем воздействии на природу антропогенного пресса. Безусловно, большое количество разнообразных и в определенной степени взаимозамещающих друг друга элементов является хорошим показателем устойчивости всей системы в целом. По–видимому, показатели экологического разнообразия могли бы также служить мерой естественной устойчивости агроценоза, препятствующей массовым размножениям вредителей.
2. БИОМАССА И ПОТОК ЭНЕРГИИ
Биомасса – это суммарная масса всех организмов данного таксона, входящих в данную экосистему. Следовательно, чтобы непосредственно определить биомассу в пределах биотопа, надо было бы собрать всех особей данного вида во всем биотопе и их взвесить. Как и в случае определения численности, это невозможно. Однако, зная численность популяции и среднюю массу особи, можно легко определить и биомассу всей популяции. В предыдущем разделе обсуждалось доминирование того или иного вида по численности. Однако в жизни биоценоза при одинаковой численности крупные объекты могут иметь гораздо большее значение, чем мелкие.
В то же время, мелкие виды в биоценозе обычно представлены большим количеством особей, чем крупные (правило М.С.Гилярова).
Взвешивание насекомых для определения их биомассы оказывается довольно сложной задачей. Выявляется, что вес отобранных из пробы насекомых довольно быстро падает сначала за счет испарения влаги, оказавшейся на покровах (особенно после использования спирта, формалина и т.д. для консервирования материала), а потом в результате испарения внутренней жидкости сквозь покровы. Обычно при переходе от одного типа испарения к другому наблюдается некоторое замедление испарения. Вес в этот момент и считается истинным показателем биомассы насекомого. Очевидно, что этот метод требует много времени и труда.
Иногда используют в качестве показателя биомассы сухой вес насекомых, но это достаточно сложно в полевых условиях. Содержание воды в теле насекомых меняется в пределах 40–85%. Особенно много воды в теле личинок двукрылых. Кроме того, может существенно меняться вес содержимого кишечника, который, например у гусениц, составляет до 60% веса всего тела. При определении биомассы обычно измеряют вес особей разного размерного класса (часто на разных стадиях развития), а затем умножают этот вес на соответствующее количество. Иногда биомассу определяют по длине тела с использованием предварительно построенных номограмм. Этот метод дает более точные результаты, если объекты обладают удлиненным телом.
Биомасса определяется ролью данного таксона в биоценозе. Как правило, наименьшей биомассой обладают паразиты и хищники, а наибольшей – сапрофаги. Интересно, что поедающие свежую зелень, насекомые–фитофаги имеют примерно такой же суммарный вес – несколько десятков килограммов на гектар – как и питающиеся той же пищей млекопитающие в естественных биоценозах. Среди сапрофагов дождевые черви обладают большей биомассой, чем насекомые. Однако и биомасса насекомых может быть значительной. Так, при переработке 1 т навоза личинками мух масса последних в субстрате может достигать 200 кг. Конечно, чем больше органических остатков содержится в детрите и почве, тем больше биомасса обитающих там членистоногих. В обычных же почвах и в лесной подстилке биомасса мелких членистоногих составляет тысячные – десятитысячные доли процента от массы всего субстрата.
 Другая важнейшая характеристика роли популяции в биоценозе – это ее участие в общем потоке энергии. Для насекомых, как и для других гетеротрофов, исходным источником энергии являются растения. Потребляемая насекомым пища может расходоваться, во-первых, на увеличение собственной биомассы (и, соответственно, энергия при этом может передаваться следующим звеньям трофической цепи) и, во-вторых, – на процесс метаболизма.
Другая важнейшая характеристика роли популяции в биоценозе – это ее участие в общем потоке энергии. Для насекомых, как и для других гетеротрофов, исходным источником энергии являются растения. Потребляемая насекомым пища может расходоваться, во-первых, на увеличение собственной биомассы (и, соответственно, энергия при этом может передаваться следующим звеньям трофической цепи) и, во-вторых, – на процесс метаболизма.
Рис.40. Зависимость потребления кислорода от массы тела у ногохвосток. Вверху – ось абсцисс в логарифмической шкале, внизу – в арифметической. Точки – результаты измерений по четырем весовым классам: 0 –0,001 мг; >0,001 –0,1 мг; >0,1 –1 мг; >1 мг (по Ю.Б.Бызовой из Ю.И.Лернова, 1975)
Первая часть потока энергии может быть учтена путем определения калорийности тканей насекомых. Вторую часть учитывают по интенсивности потребления кислорода. При этом иногда пользуются номограммами, показывающими зависимость потребления кислорода от массы тела. Здесь четко проявляется правило, упомянутое выше: интенсивность обмена на единицу массы падает с увеличением массы животного (рис.40). При определении уровня метаболизма следует поддерживать ту же среднюю температуру, которая имеет место в тех микробиотопах, где обитает данный вид.
Ю.Одум (1975) приводит замечательный пример изменений в течение сезона всех трех характеристик популяции: численности, биомассы и потока энергии на примере прямокрылого Orchelinum fidicinum , обитающего на засоленных болотах (рис.41). Эти растительноядные насекомые имеют одну генерацию за год. Численность и биомассу определяли через каждые 3–4 дня. Соответственно, можно было подсчитать прирост массы в калориях. В лаборатории измеряли также интенсивность дыхания насекомых на разных стадиях развития. Эти данные переводили в калории, а также корректировали соответственно реальной температуре в биотопе.
Конечно, численность была максимальной в самом начале сезона после выхода из яиц личинок первого возраста, далее она уменьшалась. Суммарная биомасса, наоборот, увеличивалась с ростом насекомых. Максимальный же поток энергии, имел место перед серединой летнего сезона, когда происходило интенсивное питание нимф.
Только комплекс этих трех характеристик дает полное представление о роли вида в биоценозе. Так, Ю.И.Чернов (1975) отмечает, что при зарастании грунта в тундре интенсивность дыхания ногохвосток наиболее высока в начале сезона, когда больше мелких особей и сильнее прогрев грунта солнечными лучами.
3. ЭКОЛОГИЧЕСКИЕ НИШИ И ЖИЗНЕННЫЕ ФОРМЫ
Экологические ниши
Очевидно, что особи каждого вида не занимают биотоп в целом, а приурочены к определенным его частям. Равным образом, каждый вид занимает определенное место в цепях питания, активен в течение определенного сезона и времени суток, может существовать лишь в определенном диапазоне температуры и влажности и т.д. Если совместить друг с другом все эти ограничения, мы получим экологическую характеристику вида или, как обычно говорят, его экологическую нишу. Очевидно, что экологическая ниша многомерна и представляет собой функциональную единицу биоценоза как единой системы. Образно говоря, экологическая ниша – это "рабочее место" вида в биоценозе, которое определяет функциональный статус организма в сообществе.
 Естественно, что каждый вид, отличаясь морфологически и физиологически от других видов, в принципе, должен иметь специфическую, присущую только ему многомерную нишу. Однако в природе полная ниша никогда не реализуется, не только из-за ограниченного сочетания абиотических факторов в данном биотопе, но прежде всего из–за конкуренции между близкими видами. Экологические ниши видов никогда полностью не совпадают, но они могут значительно перекрываться по многим параметрам. Поэтому различают фундаментальную нишу – то, что в принципе, могут организмы данного вида,– и меньшую реализованную нишу, часть фундаментальной ниши, которую вид занимает, будучи тесним с разных сторон конкурентами.
Естественно, что каждый вид, отличаясь морфологически и физиологически от других видов, в принципе, должен иметь специфическую, присущую только ему многомерную нишу. Однако в природе полная ниша никогда не реализуется, не только из-за ограниченного сочетания абиотических факторов в данном биотопе, но прежде всего из–за конкуренции между близкими видами. Экологические ниши видов никогда полностью не совпадают, но они могут значительно перекрываться по многим параметрам. Поэтому различают фундаментальную нишу – то, что в принципе, могут организмы данного вида,– и меньшую реализованную нишу, часть фундаментальной ниши, которую вид занимает, будучи тесним с разных сторон конкурентами.
Соответственно, при ослаблении или уничтожении вида конкурента данный вид может проявить часто неожиданные для нас возможности. То же происходит при появлении новых для данной местности растений, когда начинают формироваться новые консорции.
Рис.41. Численность (А), биомасса (сухой вес) (Б) и поток энергии (скорость ассимиляции) (В) на 1м  в популяции прямокрылого Orchelinum fidicinum в течение сезона (по Ю.Одуму, 1975)
в популяции прямокрылого Orchelinum fidicinum в течение сезона (по Ю.Одуму, 1975)
Следовательно, в зависимости от условий один и тот же вид может иметь разные реализованные ниши. В отличие от фундаментальных, реализованные ниши, занятые в разных географических районах разными видами насекомых, могут даже совпадать друг с другом. Естественно, что эти виды, оказавшись в одинаковых условиях, выполняя одинаковые функции в биоценозе и подвергаясь сходному отбору, становятся похожими друг на друга. Крайние случаи такого сходства называют экологическим викариатом.
Какие же "рабочие места" создаются в биоценозе? По трофическим связям пищевых цепей все организмы делятся на продуцентов, производящих органическое вещество, консументов, его потребляющих, и редуцентов, разлагающих мертвые организмы, их отмершие части или выделения на составные минеральные элементы. Консументы могут быть подразделены на фитофагов, хищников и паразитов. Среди редуцентов могут быть некрофаги, уничтожающие трупы, копрофаги, потребляющие экскременты, детритофаги, поедающие мелкие органические, преимущественно растительные частицы.
Отметим, что определение трофических связей – не всегда простая задача. Во многих случаях остается неясным, являются ли пищей детритофага растительные остатки или же он питается, например, мицелием грибов, пронизывающим лесную подстилку. Не менее сложная задача – изучение питания хищников в естественных условиях. Для этого применяются довольно сложные методики, в частности серологический метод (И.И.Соболева - Докучаева, 1975).
Но экологические ниши могут быть определены и в другой плоскости – по пространственному распределению. Так, на дне водоема обитает бентос, в толще воды взвешен планктон и активно движется нектон, у поверхностной пленки обитает нейстон. В почве живут геобионты, на ее поверхности – эпигеобионты, в траве – хортобионты, на кустах – тамнобионты, на деревьях – дендробионты. Формы, обитающие преимущественно в воздушной толще, называют аэробионтами.
Третья возможная экологическая классификация – по суточной ритмике, т.е. по времени активности и ее продолжительности.
Мы привели здесь сильно урезанное подразделение экологических ниш только по трем категориям признаков, которые очевидны и часто используются биологами. Сочетание этих подразделений создает уже несколько десятков экологических группировок, вхождение в которые требует от животного ряда адаптации. Однако приспосабливаться к жизни в большинстве этих группировок можно по-разному. Можно летать с помощью крыльев, активно перемещаясь в воздухе, как большинство крупных летающих насекомых. Можно летать, хотя и тоже с помощью крыльев, но используя при этом практически только токи воздуха подобно тлям (воздушный планктон). Наконец, полет возможен с помощью паутинки, как это делают гусеницы непарного шелкопряда первого возраста, а также пауки.
Следовательно, разные способы адаптации связаны с исходными морфологическими признаками объекта: размерами, отсутствием или наличием крыльев и т.д.
Жизненные формы
Сочетание экологической адаптивной характеристики с рядом морфологических признаков приводит нас к представлению о жизненной форме. Первое определение жизненных форм было разработано еще в начале девятнадцатого века, когда крупнейший естествоиспытатель Александр Гумбольдт создал первую классификацию экологических типов растений, основываясь на конвергентном подобии, возникающем благодаря сходной роли в биоценозах различных ландшафтов. Естественно, что при таком подходе растения, принадлежащие к разным, часто далеким друг от друга систематическим группам, оказываются объединенными в одну жизненную форму. Так, к форме кактусообразных А.Гумбольдт причислил также удивительно на них похожие африканские молочаи. В дальнейшем выделением и анализом таких жизненных форм занимались многие ботаники и зоологи.
По наиболее современному определению, данному И.Х.Шаровой (1987), "жизненная форма – это сходная морфоэкологическая организация (габитус) группы организмов на любой фазе жизненного цикла с разной степенью родства, отражающая характерные черты их образа жизни в определенной экосистеме и возникающая в результате параллельной и конвергентной эволюции под влиянием сходных факторов естественного отбора". Короче говоря, жизненная форма – это приспособительный тип организмов, обладающих внешним сходством. Понятия "экологическая ниша" и "жизненная форма" отличаются друг от друга как "рабочее место" и "реально работающий на этом месте". Как писал Д.Н.Кашкаров (1944),"не видовой состав, а преобладание и соотношение тех или иных жизненных форм характеризует биоценоз".
Наиболее часто применяется иерархический тип классификации жизненных форм, в которых имеется несколько рангов подразделений. Как правило, порядок этих рангов выбирается произвольно, в зависимости оттого, что считает автор классификации более существенным.
Общая классификация для всех насекомых была предложена В.В.Яхонтовым (1969). В ней за основной критерий принято местообитание насекомых. Он различает 7 основных категорий жизненных форм, каждая из которых в свою очередь делится соответственно специализации вида. Перечислим эти основные категории: 1) геобионты – обитатели почвы; 2)эпигеобионты – насекомые, живущие на поверхности почвы; 3) герпетобионты – насекомые, живущие среди растительных и других органических остатков на поверхности почвы (очевидно, что насекомые, относящиеся к этой категории, являются или геобионтами, или эпигеобионтами), 4) хортобионты – обитатели травяного покрова; 5) тамнобионты – обитатели кустарников и дендробионты – обитатели деревьев могут рассматриваться из-за сходства приспособлений как одна жизненная форма; 6) ксилобионты – обитатели отмершей древесины; 7) гидробионты – водные насекомые.
В пределах каждой категории В.В.Яхонтов различает равноценные по рангу жизненные формы. Так, среди эпигеобионтов он выделяет, в частности, псаммобионтов, живущих на поверхности песка, петробионтов, заселяющих каменистые участки, и галобионтов – обитателей засоленных участков. Среди хорто–, тамно– и дендробионтов различаются эндо– и эктобионты, т.е. насекомые, живущие внутри растительных тканей или на поверхности растений. Таким образом, в данной классификации используются только два ранга признаков.
Классификации жизненных форм разработаны для многих отрядов или семейств. Подобные системы имеются для поденок (О.А.Чернова, 1952), личинок стрекоз (Л.Н.Притыкина, 1965), саранчовых (Ф.Н. Правдин, 1971; М.Е. Черняховский, 1971; И.В.Стебаев, 1970), ногохвосток (С.К.Стебаева, 1970), жуков–чернотелок (Г.С. Медведев, 1970), муравьев (К.В.Арнольди, 1937), личинок и имаго комаров–галлиц (Б.М. Мамаев, 1963), личинок двукрылых (Н.П.Кривошеина, 1959), личинок и имаго жужелиц (И.Х.Шарова, 1981) и ряда других групп насекомых. В ряде случаев классифицируют насекомых, не принадлежащих к какому-либо одному таксону, но объединенных местообитанием или питанием. Так, О.А.Катаев (1983) предложил экологическую классификацию насекомых, обитающих в стволах деревьев или на их поверхности.
Как правило, в этих классификациях используются подразделения нескольких рангов. Например, при составлении экологической классификации имаго жужелиц И.Х.Шарова (1981) различает до четырех рангов. Так, жужелицы типа Calathus по этой классификации относятся к классу "зоофагов", подклассу "стратобиос", серии "стратобионты–скважники" и группе "подстилочных".
Дата добавления: 2015-05-16; просмотров: 995;