Половой состав популяции
Под половой структурой популяции понимают численное соотношение самцов и самок. Практически все виды насекомых в своей основе бисексуальны, т.е. представлены самцами и самками, а в случае партеногенеза – только самками. Лишь среди некоторых термитофильных мух был отмечен гермафродитизм. Для насекомых, как и для большинства других животных типично соотношение полов близкое к 1 : 1. Однако это соотношение во многих случаях нарушается. Среди насекомых довольно широко распространен партеногенез, при котором популяция состоит только из самок или же соотношение численности полов меняется в широких пределах.
Помимо генетических причин, соотношение полов может определяться соотношением выживаемости самцов и самок, которая может быть различной. Так, в начале вспышки размножения в популяции нередко преобладают самки, а после нее иногда оказывается значительно больше самцов, чем самок. Возможно, что последнее связано с меньшими размерами самцов, которым требуется меньше корма для завершения развития (А.З. Злотин, 1989).
Иногда наблюдаются и противоположные случаи. Так, в нашей лаборатории погибла культура кожеедов Trogoderma glabrum Herbst, которым дали в качестве дополнительного корма кусочки яблок, по-видимому, содержащие остаточные инсектициды. В результате пострадали только самцы, которые оказались чувствительнее самок, а оставшиеся самки, не будучи способными к партеногенезу, погибли, не оставив потомства.
Следовательно, соотношение полов в популяции может быть хорошей характеристикой ее состояния, однако определить реальное соотношение полов в потомстве часто довольно сложно по следующим причинам.
Во-первых, соотношение полов в потомстве одной самки может зависеть от возраста последней. Соответственно, если самки начинают размножаться более или менее синхронно, соотношение полов в популяции будет определенным образом меняться во времени.
Во-вторых, при определении соотношения полов следует учитывать, что самцы могут выходить из куколок на несколько дней раньше самок (многие бабочки).
В-третьих, эффективность методов учета может оказаться различной для самцов и самок из-за различий в их поведении. Нередко особи одного пола менее заметны или более осторожны. Иногда особи разных полов располагаются, например, на разной высоте ствола, предпочитают различные уровни освещенности или влажности. В лаборатории, когда берется определенная группа особей, чтобы определить соотношение полов, не исключены ошибки из-за того, что к выходу из садка или пробирки подходят прежде всего представители того пола, которые отличаются большей подвижностью или более четко реагируют на силу тяжести.
Вариации в соотношении полов имеют большое экологическое значение. В популяциях, находящихся в состоянии депрессии, целесообразно выживание прежде всего именно самцов, несущих ту же часть генофонда, что и самки, но требующих для своего развития меньше питания. Кроме того, самцы обычно отличаются большей способностью к миграциям и, следовательно, имеют возможность найти более благоприятные условия.
В случае арренотокии (один из типов факультативного партеногенеза, когда из неоплодотворенных яиц выводятся только самцы), типичной для многих перепончатокрылых, возникает также и обратная связь. Чем меньше самцов, тем больше неоплодотворенных самок и, следовательно, тем больше будет самцов в следующем поколении (см. следующий раздел). Соотношение полов в этом случае будет зависеть, с одной стороны, от наличия самцов родительского поколения и их активности, с другой – от поведения самок, которые и после спаривания могут откладывать неоплодотворенные яйца. С арренотокией связаны также сезонные изменения соотношения полов. Так, у перепончатокрылого Encyrtus infidus Rossi соотношение полов весной близко к 1:1, а летом преобладают самки (2,7:1).
Партеногенез
Партеногенез – это размножение неоплодотворенных самок. Возможно несколько типов партеногенеза. В случае телитокии самки производят только самок и, соответственно, популяция состоит только из них. Иногда телитокия сменяется во времени половым размножением. Перед этим неоплодотворенные самки производят как самок, так и самцов. Наконец, возможен такой тип размножения, когда самцы появляются в результате партеногенетического размножения и имеют гаплоидный набор хромосом, а самки отрождаются только из оплодотворенных яиц и диплоидны. Такой тип размножения, как отмечалось выше, называют арренотокией.
В принципе, бисексуальность (амфимиксис) дает большие преимущества организму. При половом размножении больше возможностей для формирования наиболее соответствующих условиям фенотипов, возникают повышающие жизнеспособность явления гетерозиса, нейтрализуются вредные мутации, накапливается разнообразие генофонда под защитой доминантных генов. Поэтому, казалось бы, более сложное и требующее больших затрат энергии и времени бисексуальное размножение сохраняется почти во всех таксонах животных. Однако и партеногенез дает серьезные экологические преимущества. Во-первых, при партеногенезе более, чем вдвое, ускоряется размножение, так как в нем участвуют все имеющиеся в популяции особи и не тратится энергия и время на поиск полового партнера. Во-вторых, при партеногенезе имеет место генетическая изоляция, что приводит к быстрому отбору мутантов, независимо от доминантности признаков. Наконец, при партеногенезе возможно основание новой популяции только одной особью, что дает большие преимущества при заселении новых, особенно островных территорий (М.С.Гиляров, 1982).
Соответственно, партеногенез возникает или в тех случаях, когда при слабой защищенности особей и их большой смертности необходимо быстро увеличить численность популяции в природе (тли 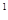 ) или же на границах ареалов, где вид редок и встреча особей разного пола затруднена. Партеногенез типичен также для относительно малоподвижных насекомых или насекомых, не способных к активному полету. Так, у мельчайших наездников-яйцеедов рода Trichogramma имеет место типичная для перепончатокрылых арренотокия, т.е. партеногенетическое отрождение самцов. Поиск брачного партнера для них затруднителен, так как зараженные трихограммой яйца хозяина встречаются относительно редко, срок жизни имаго не превышает нескольких дней, а активный полет практически не возможен из-за токов воздуха. Соответственно, вышедшие из зараженной кладки самцы трихограммы обычно на той же кладке дожидаются сестер и спариваются с ними (инбридинг). Однако благодаря арренотокии даже неоплодотворенная самка способна передать свой генофонд следующим поколениям. У некоторых же видов трихограммы типична телитокия и самцы вообще отсутствуют.
) или же на границах ареалов, где вид редок и встреча особей разного пола затруднена. Партеногенез типичен также для относительно малоподвижных насекомых или насекомых, не способных к активному полету. Так, у мельчайших наездников-яйцеедов рода Trichogramma имеет место типичная для перепончатокрылых арренотокия, т.е. партеногенетическое отрождение самцов. Поиск брачного партнера для них затруднителен, так как зараженные трихограммой яйца хозяина встречаются относительно редко, срок жизни имаго не превышает нескольких дней, а активный полет практически не возможен из-за токов воздуха. Соответственно, вышедшие из зараженной кладки самцы трихограммы обычно на той же кладке дожидаются сестер и спариваются с ними (инбридинг). Однако благодаря арренотокии даже неоплодотворенная самка способна передать свой генофонд следующим поколениям. У некоторых же видов трихограммы типична телитокия и самцы вообще отсутствуют.
Нередко партеногенез совпадает с отсутствием или недоразвитием крыльев у имаго (ряд жуков: стафилинид, короедов; некоторые бабочки: Psychidae – М.С.Гиляров, 1982). Но он возможен даже у таких хорошо летающих бабочек как тополевый бражник Amorpha populi L. Отсутствие крыльев также не обязательно влечет за собой партеногенез. Так, у известных бескрылых кузнечиков рода Saga – вид S.pedo Pall, представлен только самками, а S. ephipigerra и самками, и самцами.
Наличие партеногенетического размножения у вида не исключает и возможность полового размножения. Среди потомства партеногенетически размножающихся самок палочника Carausius morosus Br. изредка оказываются и самцы (примерно 1 самец на 1000 самок). У многих насекомых партеногенез встречается только в отдельных частях ареала, чаще всего на его краях (некоторые тараканы, богомолы, палочники, саранчовые, эмбии, жуки–долгоносики).
Иногда партеногенетическое размножение в определенное время года заменяется бисексуальным (циклический партеногенез). Включающийся половой процесс препятствует в этом случае распространению вредных мутаций. Такой циклический партеногенез имеет место у орехотворок, где партеногенетические самки и половое поколение настолько отличаются друг от друга по морфологии, что их раньше относили к разным родам. Но наиболее типичен такой циклический партеногенез для тлей. Из оплодотворенного зимующего яйца выходит самка–основательница. Эта самка дает начало многим партеногенетическим поколениям. Тли этих поколений в основном бескрылы, однако при повышенной скученности колонии на растении и плохом питании появляются отдельные партеногенетические крылатые тли, заселяющие новые растения. В конце лета под влиянием короткого фотопериода партеногенетические самки–полоноски производят самцов и самок (смена телитокии на амфитокию) и каждая оплодотворенная самка откладывает одно зимующее яйцо. Тли–полоноски обычно бывают крылатыми, иногда крылаты и самцы тлей. Появление полового поколения приурочено к концу лета, т.е. к времени расцвета популяции, когда существенно облегчается встреча полов. По-видимому, значительная часть зимующих яиц погибает, весной численность тлей низка и половой процесс в это время года был бы очень затруднен для этих малозащищенных насекомых.
У тлей наблюдаются переходы от циклического партеногенеза к постоянному. Такие изменения связаны обычно либо с выпадением из местной флоры растения, на котором поселяется самка–полоноска (сезонная смена хозяев типична для тлей), либо с заселением тлями нового района, где это растение отсутствует (А.К.Мордвилко, 1924). Естественно, что постоянный партеногенез типичен для тлей, обитающих на растениях в закрытом грунте.
У тлей скорость размножения еще более увеличивается благодаря живорождению.
Ту же цель быстрого увеличения численности и освоения субстрата преследует особый вид партеногенеза – педогенез, открытый русским зоологом Н.Вагнером (1862) у мельчайших комаров–галлиц (Itonididae). Эти комары, подобно трихограмме, мало способны к активному полету и тоже относительно мало плодовиты. Из их яиц выходят личинки, питающиеся гниющей древесиной. По мере роста личинок в них развиваются яичники. Без какого–либо оплодотворения в яичниках оказывается от 7 до 30 личинок следующего поколения, которые затем поедают изнутри тело материнской личинки и далее живут свободно. Такое педогенетическое размножение продолжается в неограниченном числе поколений, но при неблагоприятных температуре и влажности, когда возникает необходимость поиска нового субстрата, педогенез прекращается, личинки окукливаются и появляются взрослые самки и самцы.
Полиэмбрионию, т.е. появление большого количества зародышей из одной зиготы, нельзя назвать партеногенезом, хотя экологический смысл этого явления тот же – быстрое увеличение численности. Среди насекомых полиэмбриония встречается у паразитических перепончатокрылых, где одно яйцо способно сразу дать жизнь иногда нескольким тысячам личинок. Очевидно, что такое деление зародыша возможно только в очень благоприятной для развития среде, какой является внутренность тела хозяина.
5. ДИНАМИКА ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ
Динамика численности – это изменения численности популяции во времени. Эти изменения могут быть связаны с процессами, спонтанно протекающими внутри самой популяции, вызваны воздействием абиотических факторов среды или же взаимодействиями между популяциями разных видов в пределах биоценоза. Поэтому настоящий раздел учебника может считаться переходным от экологии популяций к экологии биоценозов (экосистем).
Изучая динамику численности насекомых, мы должны проводить учеты численности (плотности популяций) одновременно всех насекомых данного вида на всех стадиях их развития или же только на какой–то одной стадии. При учете численности, особенно одной стадии, будут очень четко выражены ее сезонные изменения. Так, неблагоприятный сезон насекомое обычно переживает на какой–либо одной, чаще всего покоящейся стадии развития (яйцо, куколка). В это время численность особей на других стадиях развития, как правило, равна нулю.
В течение года пики численности проявляются соответственно числу генераций, если же этих генераций много, развитие насекомых разных поколений, как правило, перекрывается. В ряде случаев длительная жизнь насекомого на какой–либо из стадий также сглаживает пики численности. Таковы, например, многие жужелицы, имаго которых живут несколько лет.
В этой главе мы уделим основное внимание изменениям численности особей какой-либо стадии данного вида или всего комплекса в течение года или же от года к году. Именно такие изменения принято называть динамикой численности. При этом следует иметь в виду, что, хотя и есть определенная корреляция между численностью на последовательных стадиях, в цикле развития она относительна и ограничивается только тем, что численность на каждой стадии развития, начиная с яйца, не должна быть больше предыдущей. Строго говоря, и это правило выполняется не всегда, так как численность популяции может увеличиваться за счет мигрантов. Соответственно, численность имаго в данном районе может оказаться много выше имевшей место численности куколок.
Многолетние наблюдения за популяциями разных видов насекомых показывают, что численность насекомых в природе из года в год различна, но эти изменения происходят в определенных пределах. Верхний предел, конечно, обусловлен имеющимися ресурсами существования данной популяции, емкостью ее среды. Нижний предел – нулевая линия, при достижении которой популяция полностью вымирает. Вполне возможно, что последнее – распространенный случай, но это не означает, что в данном биотопе на следующий год эти насекомые будут полностью отсутствовать. Иммигранты из соседних выживших популяций создадут популяцию заново.
В принципе, способность насекомых, как и других организмов, увеличивать численность популяции за счет размножения безгранична. В природе, однако, верхний предел численности практически никогда не достигается по следующим причинам.
Во-первых, при благоприятных условиях происходят спонтанные изменения генетической структуры популяции, приводящие к тому, что способность популяции к росту постепенно снижается (внутреннее сопротивление). Дело в том, что именно при благоприятных условиях выживают и дают потомство генетически неполноценные особи. В результате уменьшаются и жизнеспособность популяции в целом и ее способность к размножению. Интересно, что при определенных условиях спонтанно возникают ритмические изменения средних характеристик популяции с периодом в 1 – 2 или более поколений. По–видимому, генетические изменения свойств популяции играют большую роль в динамике численности ("волны жизни"). К сожалению, этот вопрос остается мало изученным. К этому следует добавить, что подобным же образом может меняться во времени генетическая структура популяций других организмов, взаимодействующих с данным видом: микроорганизмов, растений, других насекомых и т.д.
Во-вторых, внешняя среда, включающая в себя массу абиотических и биотических факторов, препятствует беспредельному росту численности популяции (сопротивление среды). Каждый из факторов оказывает как специфическое, так и опосредованное воздействие.
В природе можно наблюдать популяции насекомых, сохраняющиеся на протяжении десятков и сотен лет. Поэтому в литературе часто высказывается представление о колебаниях численности популяции в биоценозе как о саморегулирующемся процессе. Образно говоря, популяция рассматривается как эластичная натянутая нить, которая может быть отклонена внешними факторами до определенных пределов вверх или вниз, но при ослаблении воздействия опять возвращается к прежнему уровню.
Анализ взаимодействий популяции с окружающей средой позволяет создавать математические модели динамики численности, необходимые для прогнозирования и управления биоценозами. При построении таких моделей используют данные таблиц выживания. Очевидно, что помимо генетических спонтанных процессов уровень численности популяции должен зависеть от количественных и качественных характеристик биоценоза, от источников пищи, от численности хищников и паразитов. Влияние всех этих элементов биоценоза модифицируется реальными микроклиматическими условиями, в которых обитают или переживают неблагоприятный сезон насекомые. Конечно, микроклимат оказывает и существенное непосредственное воздействие на наблюдаемую популяцию.
При изучении динамики численности популяции некоторыми элементами, существенными для биоценоза в целом, можно пренебречь, поскольку их воздействие на эту популяцию опосредовано. Всю совокупность существенных связей и взаимодействий данной популяции называют ее жизненной системой (L.R. Clark et al., 1967). Жизненная система – это "экосистема, рассматриваемая в аспекте определенной популяции" (А.А. Шаров, 1989). Анализ взаимодействий в пределах жизненной системы позволяет понять закономерности колебаний численности насекомых. Эти закономерности имеют также большое значение для практики, так как позволяют прогнозировать изменения численности вредных и полезных насекомых, а также результаты воздействия различных искусственных биологических и химических агентов.
Биотический потенциал насекомых
Предположим, что самка данного вида откладывает в среднем 200 яиц (плодовитость F равна 200) и смертность на протяжении всего развития равна нулю. Если соотношение полов в потомстве, как чаще всего бывает 1:1 (доля самок q= 0,5), то это означает, что в первом поколении будет Fq т.е. 200 0,5 = 100 самок. Каждая из этих самок в следующем поколении даст жизнь еще сотне самок, в результате чего во втором поколении будет 10000 самок. Очевидно, что в n–ном поколении число самок может быть рассчитано по следующей формуле:
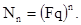
Если же исходно мы имеем не одну самку, а N самок, то через n поколений их будет:
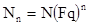 (1)
(1)
Очевидно, что при таких условиях численность популяции будет круто возрастать по экспоненте (степенной функции). Смена поколений все же занимает некоторое время. Тогда скорость изменения численности при большом количестве поколений или их быстрой смене можно представить как результат деления прироста численности 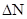 на интервал времени
на интервал времени 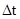 (абсолютная скорость роста популяции), или из расчета на исходное число особей
(абсолютная скорость роста популяции), или из расчета на исходное число особей  -
-
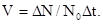
При последовательном уменьшении интервала времени ( 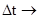 0) мы получаем мгновенную скорость роста популяции – r (биотический потенциал):
0) мы получаем мгновенную скорость роста популяции – r (биотический потенциал):
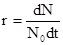 (2)
(2)
Возвращаясь к формуле роста популяции (1), мы можем теперь ее написать следующим образом:
 (3)
(3)
где 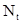 – численность популяции через время t, N
– численность популяции через время t, N 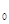 – исходная численность популяции, е – основание натуральных логарифмов, r – биотический потенциал, t – интервал времени. График этой экспоненциальной (показательной) функции представлен на рис.27. Если логарифмировать формулу 3, получим следующее выражение:
– исходная численность популяции, е – основание натуральных логарифмов, r – биотический потенциал, t – интервал времени. График этой экспоненциальной (показательной) функции представлен на рис.27. Если логарифмировать формулу 3, получим следующее выражение:
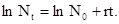 (4)
(4)
График этой функции – прямая линия. Биотический потенциал на этом графике может быть представлен как тангенс угла наклона графика к оси абсцисс. Очевидно, что биотический потенциал – не чисто умозрительная категория. Зная численность популяции N  в момент времени t
в момент времени t 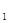 , и последующую численность N
, и последующую численность N 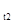 в момент t
в момент t 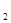 , можно определить биотический потенциал по формуле:
, можно определить биотический потенциал по формуле:
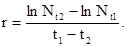 (5)
(5)
В начале мы приняли, что смертность насекомых в течение развития равна нулю. При такой ситуации биотический потенциал будет максимально возможным в данных условиях. В природе же это условие практически никогда не выполняется и определяемый биотический потенциал будет определяться разностью между плодовитостью и смертностью. Благодаря стремлению размножаться, насекомые могли бы увеличивать свою численность беспредельно, если бы не тормозящие рост популяции факторы, снижающие плодовитость или приводящие часть насекомых к гибели. Такое сопротивление среды можно определить как разность между максимально возможным и реально наблюдаемым биотическим потенциалом .
Насекомые, будучи мелкими существами отличаются исключительно высоким биотическим потенциалом. Среди них зарегистрированы величины этого потенциала 0,5–0,8 (недельный прирост популяции), а у тлей – даже 0,3 в сутки. Наиболее высокий среди позвоночных животных потенциал отмечен также у самых мелких форм – грызунов. Однако недельный потенциал у этих животных не превышает 0,1.
Естественно, что у насекомых r–стратегов биотический потенциал всегда выше, чем у К–стратегов, которые приспособлены к существованию в пределах узкой экологической ниши. При высоком биотическом потенциале К–стратеги разрушили бы необходимый для их существования биоценоз.
Подчеркнем, что высокое значение биотического потенциала означает возможность возникновения резких вспышек численности, опасных для хозяйственной деятельности человека. Кроме того, способность быстро увеличивать свою численность является основой для использования насекомых как источника животного белка. Скорость наращивания биомассы у насекомых гораздо выше, чем у каких–либо других животных, за исключением, пожалуй, ракообразных. Насекомые могут быть белковой основой для корма домашней птицы, для рыб и, в какой–то мере, для скота. Добавим, что в странах Юго-Восточной Азии насекомые являются обычной пищей человека. Поэтому разведение насекомых для кормов, а может быть, и пищи является одной из перспектив обеспечить жизнь быстро растущего человечества.

Рис.27. Рост численности популяции без ограничивающих воздействий. Показан рост численности одной и той же гипотетической популяции в арифметическом (слева) и логарифмическом (справа) масштабе (по Ю.Одуму, 1986)
Роль абиотических факторов среды.
Влияние абиотических факторов на насекомых подробно рассмотрено в предыдущих главах. Совершенно очевидно, что эти факторы, существенно изменяя уровень смертности и плодовитость насекомых, могут быть одной из основных причин изменений их численности во времени.
Остановимся отдельно на воздействии на численность насекомых некоторых наиболее важных абиотических факторов.
Температура, как ниже оптимума, так и выше его, приводит к замедлению развития, а при ее экстремальных значениях – к гибели насекомых. Температура может влиять и на состояние кормового растения, увеличивая или уменьшая его ценность, а также на популяции конкурентов, хищников и паразитов.
Степень воздействия неблагоприятной температуры зависит от численности популяции. Действительно, если бы среда обитания была бы полностью однородной в отношении температуры, то ее влияние сводилось бы к гибели определенного процента особей, независимо от численности популяции.
Строго говоря, к сопротивлению среды здесь добавляются генетические изменения структуры популяции, которые при высокой численности снижают плодовитость, а также эмиграция.
Но температура даже в пределах одного растения может различаться на 5–19°, в пределах же небольшого участка травянистой растительности этот контраст может достигать 20° и выше. Чем больше численность популяции, тем труднее найти незанятое укрытие, и тем больше популяция подвержена воздействию неблагоприятных факторов.
Влажность влияет на численность популяции аналогичным образом. Здесь особняком стоят сосущие фитофаги, которые всегда имеют достаточно влаги, независимо от ее содержания в воздухе.
Катастрофические факторы могут существенно изменять численность популяций. Так, выжигание весной сухой травы приводит к гибели многих насекомых. То же можно сказать о затоплении водой. Однако в ряде случаев, если затапливание является регулярным, оно предусматривается жизненным циклом и насекомые на отдельных стадиях оказываются выносливыми по отношению к этому воздействию. Чаще же насекомые заранее мигрируют на более возвышенные участки.
Однако при наблюдениях в природе оказывается довольно сложно понять, какие из факторов среды наиболее существенно влияют на численность популяции. Так, наблюдения за численностью одного вида трипсов, проведенные на протяжении 14 лет, показали четкую связь численности популяции с погодой (J.Davidson, H.G.Andrewartha, 1948). Эти авторы, используя метод множественной регрессии, получили следующее уравнение:
log Y = –2,390 + 0,125х + 0,202 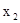 +0,186x
+0,186x 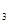 + 0,085х
+ 0,085х  ,
,
где Y – реальная численность популяции, x 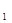 – сумма эффективных температур за сезон, x – суммарное количество осадков за два первых весенних месяца, x – средняя эффективная температура за эти же месяцы и x – значение x в предыдущий год. Учитывая данные факторы, авторы смогли объяснить до 84% колебаний численности этого вида. Подчеркнем, что этот яркий пример, показывающий статистическую связь численности популяции трипсов с погодными факторами, все же не доказывает, что именно сама погода влияет на трипсов. От погоды, несомненно, зависят состояние кормового растения, а также активность паразитов, хищников и патогенных микроорганизмов.
– сумма эффективных температур за сезон, x – суммарное количество осадков за два первых весенних месяца, x – средняя эффективная температура за эти же месяцы и x – значение x в предыдущий год. Учитывая данные факторы, авторы смогли объяснить до 84% колебаний численности этого вида. Подчеркнем, что этот яркий пример, показывающий статистическую связь численности популяции трипсов с погодными факторами, все же не доказывает, что именно сама погода влияет на трипсов. От погоды, несомненно, зависят состояние кормового растения, а также активность паразитов, хищников и патогенных микроорганизмов.
По–видимому, именно погода оказывает первичное воздействие на жизненную систему, выводя ее из равновесного состояния, приводя прямо или косвенно либо к гибели значительной части популяции, либо способствуя ее расцвету. Правда, влияние погоды зависит от исходного состояния популяции. Известно, что благоприятная погода, приходящаяся на спад численности насекомого, не в состоянии сразу же вызвать новую вспышку.
Многолетние наблюдения показывают довольно резкие изменения численности популяции от года к году как у трипсов, так и у других насекомых (рис.28). Интервалы между массовыми размножениями тех или иных насекомых зависят от вида насекомого и географической зоны. Они могут быть равны 2, 4, 5, 10, 20 и более годам, но очень часто максимумы повторяются каждые 10–14 лет (Н.С. Щербиновский, 1964; А.С.Рожков, 1965; Ю.П. Кондаков, 1974; H.Eidmann, 1931). Максимумы размножения в разных географических зонах обычно возникают независимо друг от друга. Они могут не совпадать во времени даже в разных насаждениях одного лесхоза. Такова, например, динамика численности непарного шелкопряда (В.И. Бенкевич, 1984).
В некоторых же работах описаны синхронные вспышки размножения, охватывающие одновременно очень большие территории. В частности, В.Мартинек (V.Martinek, 1968) описал подобную динамику численности рыжего соснового пилильщика на территории Евразии.
Чаще всего массовые размножения насекомых представляются связанными с погодой, а именно с засухой. После засухи отмечаются вспышки у саранчевых, непарного шелкопряда, сибирского шелкопряда. Не исключено, что засухи вызывают и массовое размножение рыжего соснового пилильщика.

Рис.28. Многолетние колебания численности двух видов бабочек. Вверху – сосновая пяденица Bupalus piniarius L. в сосновом бору близ Литцлингена (Германия); внизу – лиственничная листовертка Zeiraphera griseana Hbn. в лиственничном лесу долины Энгадин (Швейцария) (по Ю. Одуму, 1986)
Особый интерес вызывает возможная связь вспышек массового размножения насекомых с солнечной активностью. Наличие такой связи позволило бы легко прогнозировать эти вспышки, поскольку солнечная активность относительно периодична. Длительность основного цикла солнечной активности равна примерно 11–12 годам. Действительно, во многих случаях вспышки численности различных насекомых повторяются каждые 10–14 лет и пики размножения совпадают с той или иной фазой солнечной активности: с максимумом у многих бабочек, со спадом у некоторых саранчовых и рыжего соснового пилильщика, с минимумом у лугового мотылька, с началом подъема солнечной активности у мигрирующей саранчи. Повторение массовых размножений этого вида саранчи и их совпадение с определенными фазами солнечного цикла было отмечено еще в прошлом веке (Ф.Кеппен, 1870). Аналогичные работы, основанные на многолетних наблюдениях за динамикой численности тех или иных видов, появляются и в наши дни. Скорее всего, такая связь не является случайным совпадением. Здесь возможно либо непосредственное воздействие каких–то факторов, циклически изменяющихся под влиянием солнечной активности, либо так называемый "захват циклов", генерируемых по причинам, не связанным с солнечной активностью.
Согласно Б.М.Владимирскому (1980), геофизическими факторами, непосредственно зависимыми от солнечной активности и, по–видимому, способными оказывать влияние на биологические объекты, являются: электромагнитное поле Земли, особенно переменное в диапазоне 10 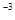 –10 Гц; инфразвуки, возникающие при полярных сияниях и распространяющиеся по всей планете; увеличение мощности относительно коротковолнового ультрафиолетового излучения в диапазоне около 290 нм в связи с изменениями озонового экрана Земли; увеличение радиоактивности атмосферы, благодаря повышению концентрации газа радона.
–10 Гц; инфразвуки, возникающие при полярных сияниях и распространяющиеся по всей планете; увеличение мощности относительно коротковолнового ультрафиолетового излучения в диапазоне около 290 нм в связи с изменениями озонового экрана Земли; увеличение радиоактивности атмосферы, благодаря повышению концентрации газа радона.
Могут ли эти факторы влиять на численность популяций? Мы отмечали выше, что резкие изменения активности насекомых нередко коррелируют с геомагнитными бурями – явлением, непосредственно связанным с солнечной активностью. Одним из основных действующих факторов здесь предполагается низкочастотное электромагнитное поле. Однако нам известно лишь одно указание на возможность влияния подобных искусственных полей на плодовитость насекомых, а следовательно, и на динамику их численности. По предварительным наблюдениям В.Б.Чернышева и В.М.Афониной(1971), такое поле влияет на плодовитость партеногенетических самок акациевой тли. Это влияние неоднозначно. В начале лета, когда происходит быстрый рост популяции тлей, поле достоверно повышает их плодовитость на 30%. В конце лета, когда численность популяции тлей на спаде, поле подавляет их размножение примерно на 25%. Между этими периодами действие поля неопределенно.
Конечно, возможно и более косвенное воздействие электромагнитного поля на динамику численности насекомых. В опытах искусственное поле существенно ускоряет рост колонии некоторых микроорганизмов. Если это так, то повышенная солнечная активность может способствовать возникновению эпизоотии среди насекомых.
Инфразвуки могут влиять на уровень поведенческих реакций насекомых, однако их роль в динамике численности не исследовалась.
Даже сравнительно небольшие изменения спектра ультрафиолетовых лучей явно могут отразиться на динамике численности насекомых. При этом возможно как непосредственное воздействие на организм насекомого, так и косвенное, связанное с бактерицидным действием этого излучения. Подобным же образом на динамику численности могут воздействовать и изменения уровня радиоактивности атмосферы как явно мутагенного фактора. Однако эти вопросы остаются практически неизученными. Нам известна лишь одна работа (Levengood, Shinkle, 1960), согласно которой повышенная солнечная активность приводит к снижению плодовитости мух дрозофил, но только находящихся в сильном магнитном поле. Авторы связывают эти изменения с тем, что магнитное поле улавливает частицы высоких энергий и тем самым усиливает эффект солнечной активности. Однако этим исследователям не удалось найти прямой корреляции величины эффекта с числами Вольфа, показывающими уровень солнечной активности.
Гораздо более вероятным представляется воздействие солнечной активности через погоду. Известно, что солнечная активность приводит к определенным изменениям циркуляционных процессов в атмосфере. Связь же массовых размножений с погодой не вызывает никаких сомнений. Конечно, погода может влиять и непосредственно на организм насекомого, и косвенно, изменяя его питание и окружение.
Многолетние циклические изменения численности насекомых, конечно, могут быть вызваны многими причинами помимо воздействия солнечной активности. Как отмечалось выше, цикличность может быть связана со спонтанными изменениями генетической структуры популяции, а также возникать в результате взаимодействия популяции с хищниками и паразитами, из-за внутривидовой конкуренции, из-за того, что под влиянием фитофага растение становится более иммунным по отношения к нему и по многим другим причинам. Таким образом, популяции, взаимодействуя со средой, способны сами генерировать колебания своей численности. При этом возможен так называемый захват автоколебаний в популяции внешней цикликой метеорологических или геофизических факторов.
Конкуренция между особями одного вида
Очевидно, что пищевые и пространственные ресурсы любого вида ограничены. Предположим, что верхний предел численности какой–то популяции равен К. Тогда формула 2 примет следующий вид:
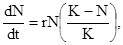
где dN/dt – скорость увеличения популяции, r – биотический потенциал, N – численность популяции в каждый предшествующий момент времени.
Сами же изменения численности в зависимости от времени описываются так называемой логистической кривой (уравнение Ферхюльста – Перла):
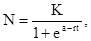
где а = 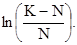
Эта кривая показана на рис.29, где также указаны точками данные эксперимента с жуками зерновыми точильщиками (Rhyzopertha dominica F.) (A.C.Crombie,1945). Этих жуков разводили в небольших объемах зерна (10 г), причем корм еженедельно просеивали, удаляя все отходы и добавляя свежее зерно. Примерно через полгода каждая такая лабораторная популяция жуков достигала устойчивого уровня (около 340 жуков).

Рис.29.Нарастание численности жуков – зерновых точильщиков Rhyzopertha dominica F. в лабораторной культуре с ограниченным количеством корма (10 г зерен пшеницы, которые каждую неделю просеивали и дополняли до 10 г). Точками показаны результаты реальных наблюдений, кривая – расчет по формуле логистической зависимости, приведенной на рисунке (по А.Кромби, из Дж. Варли и др., 1978)
Логистическая кривая все же обычно оказывается недостаточной для описания сложных процессов в популяции, происходящих при ограниченны объеме пищевых ресурсов. Если подсчитывать не только имаго, но и насекомых на всех стадиях развития, получаются более сложные кривые, часто включающие в себя и ритмические компоненты. Простейшая модель конкуренции была предложена Р.Ф.Моррисом (F.R.Morris, 1959). По его мнению, численность следующего поколения N 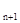 может быть связана с численностью предшествующего поколения N
может быть связана с численностью предшествующего поколения N 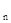 следующим образом:
следующим образом:
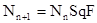 , (8)
, (8)
где S – доля выживших особей, q – доля самок в поколении n, а F – их средняя плодовитость.
Величина S q F – это коэффициент размножения. Он показывает соотношение численностей последующего и предыдущего поколений и обычно обозначается греческой буквой 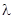 :
:
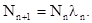
Мы употребляем здесь коэффициент размножения с символом n , поэтому что от поколения к поколению этот коэффициент может изменятся. При логарифмировании предыдущая формула будет выглядеть следующим образом:
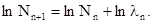
Величина 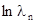 показывает изменения численности популяции, она может быть больше нуля (популяция возрастает), равна нулю (популяция стабильна) или меньше нуля (численность популяции уменьшается).
показывает изменения численности популяции, она может быть больше нуля (популяция возрастает), равна нулю (популяция стабильна) или меньше нуля (численность популяции уменьшается).
С ростом численности популяции обычно возрастает смертность, поэтму график зависимости от численности предыдущего поколения 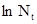 может быть приближенно описан как линейная зависимость типа
может быть приближенно описан как линейная зависимость типа
y = а – bx
или в нашем случае:
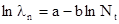
Очевидно, что b – тангенс угла наклона графика к оси абсцисс. Если угол наклона меньше 45° и график идет полого, то численность будет постепенно приближаться к устойчивой точке, где равен нулю (точка пересечения графика с осью абсцисс). Если угол наклона равен 45°, то компенсация возрастания численности будет иметь место уже во время следующего поколения. Если же тангенс угла наклона больше двух и график круто идет вниз, то в популяции должны возникнуть колебания со все возрастающей амплитудой. Следовательно, вся система устойчива, только в том случае, если 0<b<2.
Возникновение в лабораторной популяции четко выраженной цикличности, связанной с недостатком корма, было показано на примере культуры мух Lucilia cuprina (A.J.Nicholson, 1950,1954).

Рис.30. Зависимость между плотностью лабораторной популяции личинок и численностью имаго мух Lucilia cuprina. Пунктирной линией показан рост численности, который имел бы место при отсутствии внутривидовой конкуренции (по Л.Никольсону из Э.Макфедьена,1965)
В этих опытах имаго мух, находившиеся в садках, с избытком получали корм и, соответственно, откладывали много яиц. Эти яйца помещали в сосуд, где все вышедшие личинки получали одну строго ограниченную порцию пищи – 50 г мяса в день. В результате с ростом количества яиц и, соответственно, личинок, через несколько поколений наступал такой момент, когда конкуренция между личинками за пишу оказывалась настолько жесткой, что большинство из них не могло закончить развитие и погибало (рис.30). В этом поколении резко уменьшалось число взрослых мух и опять создавались благоприятные условия для развития личинок следующего поколения. Колебания численности (рис.31) возникали именно потому, что возрастание количества имаго усиливало конкуренцию не сразу же, а только в следующих поколениях. В других своих опытах этот же автор получил аналогичные колебания численности имаго, ограничивая их в пище, но при избытке корма для личинок.
Такая ситуация, когда численность популяции доходит до верхнего предела, обусловленного количеством пищи, легко достижима только в лаборатории. В природе погода, конкуренты, паразиты и хищники могут существенно сдерживать рост популяции. Кроме того, известен ряд приспособлений, препятствующих появлению жесткой конкуренции за пищу или пространство.
С ростом численности популяции обычно резко возрастает стремление особей мигрировать. При этом существенно расширяется пространство, занятое популяцией в основном за счет частей биотопа, менее благоприятных для данного вида. При этом происходит также расслоение пространственной структуры популяции. Так, в самом населенном, но наиболее благоприятном по другим условиям месте имеют больше шансов выжить менее жизнеспособные и малоплодовитые особи, не способные к дальним миграциям. Наоборот, чем дальше от центра зоны, занимаемой популяцией, тем чаще должны попадаться особи с высокими миграционными способностями и нередко с большой  плодовитостью (Г.В. Гречаный, 1990).
плодовитостью (Г.В. Гречаный, 1990).
Рис.31. Развитие культуры мух Lucilia cuprina при избытке корма для имаго (А) и при введении строгого рациона для них (Б). Личинки получали ограниченное количество корма. Сплошная кривая – число имаго, вертикальные линии – число имаго в следующем поколении, полученных из яиц, отложенных в данный день. При избытке корма имаго откладывали настолько много яиц, что большинство личинок погибали из-за конкуренции. При этом возникали также колебания численности (по Л.Никольсону из Э.Макфедьена, 1966)
В принципе, возможен и другой вариант, особенно в случае медленно меняющихся во времени биотопов и наличия резко выраженного территориального поведения. Тогда более сильные особи остаются в благоприятном по условиям центре, вытесняя оттуда более ослабленных. В этом случае перенаселение вообще невозможно, ослабленные же особи в неблагоприятных условиях имеют больше вероятности погибнуть.
В любом случае популяция как бы растекается в пространстве, не достигая слишком высокой плотности. При этом у ряда видов имеют место специальные приспособления, обеспечивающие резкое увеличение миграционных возможностей с ростом плотности популяции.
Так, у тлей при высокой скученности, как говорилось выше, рождаются крылатые партеногенетические особи, которые менее плодовиты, но способны к полету. У мигрирующей саранчи Locusta migratoria L. при большой концентрации личинок они развиваются в мигрирующую "фазу", внешне сильно отличающуюся от одиночной. До открытия этих фазовых изменений Б.П.Уваровым мигрирующую и одиночную саранчу считали разными видами.
Частые контакты между особями паразитических перепончатокрылых иногда приводят к откладке ими неоплодотворенных яиц, из которых обычно выводятся только самцы и таким образом сдерживается рост популяции. У трихограммы отмечен несколько иной механизм. При большой скученности популяции самки откладывают свои яйца в уже зараженные другими самками яйца хозяина. В результате конкуренции личинок происходит их существенная гибель, причем гибнут в первую очередь требующие больше корма личинки самок (А.Дахия, 1991).
При очень жесткой конкуренции личинки мух могут развиваться в мелких половозрелых особей, плодовитость которых существенно снижена. Возможно и появление вполне нормальных, но малоплодовитых имаго.
Широко распространенный способ автоматического снижения численности популяции при ее высокой плотности – это каннибализм. Взрослые насекомые нередко поедают яйца, личинок и куколок своего же вида (например, малый мучной хрущак Tribolium castaneum Host). Хищные личинки, ведущие одиночный образ жизни, при встрече часто поедают друг друга, что очень обычно, например, для личинок златоглазок. Однако каннибализм возможен и у типичных фитофагов. Так ведут себя, например гусеницы капустной совки. Проблема каннибализма стоит особенно остро в связи с массовым разведением насекомых. По-видимому, стремление к каннибализму может быть снижено с помощью специально составленных пищевых диет.
Помимо механических взаимодействий друг с другом, приводящих к рассредоточению популяции в пространстве или снижению плодовитости, возможны и химические взаимодействия. Так, жуки – мучные хрущаки Tribolium выделяют этилбензохинон и другие вещества, которые обладают спермицидным действием, в результате чего ограничивается рост их численности. В принципе, и загрязнение экскрементами субстрата, на котором развиваются личинки, обычно приводит к снижению жизнеспособности последних. Есть основания предполагать, что некоторые насекомые способны выделять и вещества, являющиеся репеллентами для других особей того же вида. Поиск таких веществ мог бы быть весьма перспективным как в защите растений, так и в борьбе с переносчиками заболеваний или домашними насекомыми.
Итак, по ряду причин популяции в природе лишь в очень редких случаях достигают такого уровня, при котором возникает резко выраженная внутривидовая конкуренция. Однако даже при относительно низкой численности наилучшая пища и наиболее благоприятные укрытия достаются, по–видимому, наиболее жизнеспособным насекомым. Чем больше численность популяции, тем чаше используется менее пригодная пища, неудобные и плохо защищенные местообитания.
Однако во многих случаях низкая численность оказывается также неблагоприятной для жизни насекомых, как и высокая (принцип Олли – W.C.Allee et al.,1949). Из-за привлечения особями одного вида друг друга насекомые скапливаются в местах с наиболее благоприятным микроклиматом и обильной пищей (скопления божьих коровок в местах зимовок или тараканов в укрытиях). При объединении мигрирующих насекомых в стаи достигается их большая защищенность от хищников. Кроме того, при скоплениях насекомых на определенных участках значительно облегчается встреча полов. Некоторые насекомые приспособлены к групповой жизни, когда малое количество особей не может существовать. Таковы, например, личинки мух с их групповым перевариванием пищи, гусеницы, живущие в групповых паутинных гнездах, защищающих их от пониженной влажности и врагов, личинки пилильщиков, совместно обороняющиеся при нападении. Мы не говорим здесь об общественных насекомых, так как их скопление в гнезде нельзя рассматривать как высокую плотность популяции. Однако если воспринимать колонию общественных насекомых как некий суперорганизм, то взаимоотношения между колониями будут в какой–то мере аналогичны взаимоотношениям между особями не общественных насекомых.
В конце этого раздела нельзя не упомянуть об особом виде конкуренции, возникающей между самцами за обладание самкой. Здесь также обычно выигрывают наиболее жизнеспособные и агрессивные самцы. Это особенно очевидно, если наблюдать брачные драки между самцами, типичными для некоторых дневных бабочек и жуков.
Конкуренция между видами
Одни и те же пищевые ресурсы или местообитания могут быть использованы несколькими видами. Очевидно, что при этом на внутривидовую конкуренцию за ресурсы накладывается и межвидовая. Для моделирования таких конкурентных отношений можно изменить приведенную выше формулу логистической зависимости (6) следующим образом:
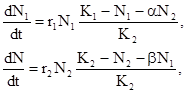
где N и N – численности этих двух видов, r и r – биотические потенциалы этих видов, K и K – пределы численностей этих видов, связанные с ограниченностью ресурсов, коэффициент 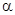 показывает степень воздействия второго вида на первый, а
показывает степень воздействия второго вида на первый, а 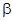 – первого вида на второй.
– первого вида на второй.
Анализ этих уравнений показывает, что здесь возможны четыре ситуации: вытеснение вида 1 видом 2; вытеснение вида 2 видом 1; неустойчивое равновесие, приводящее в конце концов к гибели одного из видов и устойчивое равновесие. Последнее возможно только в том случае, если каждый вид при увеличении своей численности подавляет сам себя больше, чем другой вид. В случае вытеснения одного вида другим исход определяется уровнем биотического потенциала и способностью выживать при недостатке пищевых ресурсов.
Т.Парк (T.Park, 1948) изучал в лаборатории конкуренцию между двумя близкими видами мучных хрущаков Tribolium castaneum Herbst. и T.confusum Dur. Первый вид лучше развивается при повышенных температуре и влажности, а второй – в прохладных и более сухих условиях. Побеждал в конкуренции всегда тот вид, жизни которого больше соответствовали условия эксперимента. Однако если условия были нестабильными и менялись во времени, оба вида сосуществовали неопределенно долгое время.
В некоторых случаях насекомые способны, подобно хищникам, поедать своих конкурентов. В опытах А.Кромби (A.Crombie, 1946) вместе поселяли личинок малого мучного хрущака Tribolium confusum Dur. и суринамского мукоеда Oryzaephilus surinamensis L. Личинки мучного хрущака охотно поедали куколок мукоеда. В том случае, если все личинки и куколки находились в муке, суринамский мукоед быстро прекращал свое существование и эксперимент заканчивался. Однако в зернах пшеницы выживали оба вида, так как в узких пространствах между зернами часть куколок мукоеда оказывалась не доступной для хрущака. Того же эффекта удавалось добиться и в муке, если в нее помещали обрезки узких стеклянных трубочек, в которых личинки мукоеда могли безопасно окуклиться.
Итак, разнородность условий во времени и пространстве создает условия для сколь угодно долгого сосуществования даже близких видов. Любые два вида отличаются друг от друга не только по морфологическим и биохимическим признакам, но также и по своим экологическим возможностям. Следовательно, в естественной среде, к которой они приспособлены, всегда найдутся место и время, когда данный вид будет процветать, несмотря на присутствие мощных конкурентов.
Блестящим примером такого сосуществования являются насекомые, развивающиеся под корой деревьев: короеды и некоторые усачи. Каждый из видов предпочитает для поселения свою зону с определенным прогревом и увлажнением на определенной высоте над уровнем земли.
То, что каждый вид занимает свою экологическую нишу, не означает, однако, что нет конкурентных взаимоотношений на пограничных участках. Так, А.Понтин (A.Pontin, 1969) изучал сосуществование колоний муравьев Lasius niger L. и Lasius flavus F. Он показал, что плодовитость в колониях обоих видов увеличивается, если удалить близлежащие колонии L.flavus. Интересно, что колонии L.flavus больше угнетались своим же видом, чем чужим. Как мы указывали выше, резко выраженная внутривидовая конкуренция является основой для устойчивого сосуществования конкурирующих видов.
Вообще, в природе взаимоотношения между видами уже настолько "притерты", что заметить какую–либо конкуренцию между ними не всегда возможно. Нарастание численности видов обычно сдерживается и другими биотическими и абиотическими факторами и далеко не всегда доходит до такого уровня, когда начинает проявляться конкуренция за ресурсы. Правда, при обитании в быстро разлагающихся запасах органической пищи, таких как трупы или экскременты, конкуренция может быть очень жесткой. Здесь взаимоотношения между видами отличаются высокой напряженностью, что приводит, с одной стороны, к выработке тончайших органов чувств у взрослых насекомых, которые позволяют находить эти запасы пищи возможно раньше, с другой – в таких кратковременных местах обитания четко выражена сукцессия, т.е. смена комплексов видов во времени.
Если же виды приспособлены приблизительно к одинаковым стадиям сукцессии, то конкуренция между ними может быть особенно суровой. Личинки мух, например, при обитании на падали или в навозе выделяют ряд химических веществ, препятствующих появлению питающихся тем же субстратом нематод и грибов.
Всем хорошо известно, что на жуках–могильщиках рода Necrophorus часто встречаются гамазоидные клещи Poecilochirus necrophori. Жуки переносят этих клещей с одного трупа на другой, обеспечивая им расселение. Но клещи оказываются исключительно полезными для могильщиков, так как уничтожают яйца падальных мух – главных конкурентов жуков–могильщиков.
Резко выражена также конкуренция за хозяина у перепончатокрылых паразитов.
Яркие примеры конкуренции дает случайная или намеренная интродукция новых видов. Так, в итоге "вторжения" рыжего таракана Blatella germanica L. в Восточную Европу ранее очень распространенный здесь черный таракан Blatta orientalis L., по–видимому, обладающий меньшим биотическим потенциалом, был вытеснен почти из всех мест своего обитания и сохранился в настоящее время преимущественно в подвальных помещениях около водопроводных труб.
Другим примером может служить интродукция на Гавайские острова паразитов для борьбы с плодовой мухой Dacus dorsalis. Три вида наездников–браконид, завезенные на эти острова, последовательно вытесняли друг друга (H.A.Bess et ai., 1961).
Взаимодействия насекомого–фитофага и растения
В случае повреждения насекомыми растение способно "мобилизовать" защитные реакции и повысить свой иммунитет. При этом растение изменяет определенным образом свой химический состав, и начинает отпугивать насекомых. Вырабатываемые растением вещества могут препятствовать усвоению корма (например, при появлении в тканях растения ингибиторов протеиназы) или снижать плодовитость насекомых. В некоторых же случаях растение становится токсичным для фитофага. Широко распространены и другие способы самозащиты растения от поедающего его насекомого, например выделение смолы, в которой гибнет насекомое, или появление коркового слоя, мешающего питанию. Сложная картина механизмов, обеспечивающих устойчивость растения к насекомым подробно описана в работе И.Д.Шапиро и Н.А.Вилковой (1973). Таким образом, растение способно в определенной степени сдерживать увеличение популяции фитофага.
Однако при массовом размножении фитофага растение ослабляется и становится неспособным противостоять нападению. В этом случае возникает положительная обратная связь, приводящая в конце концов к распаду системы, т.е. к гибели растения. Интересный пример таких взаимодействий приводят А.С.Исаев и др. (1984), описывая динамику взаимодействия большого хвойного черного усача (Monochamus urussovi Fisch.) с его кормовым деревом – сибирской пихтой. Личинки этих жуков питаются лубом и частично древесиной, пихта же отлично защищается от них, интенсивно выделяя смолу. Но только здоровые деревья способны противостоять нападению усача. Поэтому жуки развиваются в основном за счет ветровала и отдельных старых и ослабленных деревьев, не причиняя вреда лесу. В какой-то мере жуки могут быть даже полезными, устраняя отстающие в росте деревья. Имаго же питаются, объедая кору с веток и иногда даже их окольцовывая. В норме этих жуков немного и они не наносят деревьям существенного вреда. Такая устойчивая популяция усача может существовать сколь угодно долго.
Совсем иная ситуация возникает при ослаблении и гибели части деревьев под влиянием пожаров, промышленных выбросов или после массового размножения сибирского шелкопряда, объедающего всю хвою. В этом случае жуки исходно имеют много субстрата, пригодного для развития личинок. Жуки размножаются в массе, и повреждений, наносимых имаго, становится так много, что от них ослабляются и практически здоровые деревья. Численность популяции резко возрастает, и вспышка кончается только после гибели всех пихт в лесу.
В экологической литературе хищникам и особенно паразитам уделяется большое внимание, так как предполагается, что именно они регулируют численность популяции, увеличивая свой пресс при массовом размножении их жертв и ослабляя его в годы с низкой численностью.
Здесь мы основное внимание уделим паразитам, как влияющему на численность фактору. Модели хищничества очень близки к моделям паразитизма. Основное различие заключается в том, что у паразита число отложенных им яиц определенным образом соответствует числу найденных им хозяев, у хищника же эта связь не столь однозначна.
Простейшая модель взаимодействия популяций хозяина и паразита была представлена У.Томпсоном (W.R.Thompson, 1924). Эта модель была разработана для случая, когда плотность популяции хозяина исходно высока и поэтому паразит тратит мало времени на поиск. Предполагается также, что при каждой встрече самка паразита будет откладывать в хозяина по одному яйцу, (что не исключает возможность повторного заражения при последующей встрече). Тогда число отложенных всеми паразитами яиц N (=число встреч с хозяином) может быть определено по формуле:
N = PF,
где P – численность (плотность) паразита, а F – его средняя плодовитость. Если плотность определяется в расчете на 1 м 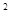 , то PF – это количество яиц, отложенных на 1 м .
, то PF – это количество яиц, отложенных на 1 м .
При случайном (пуассоновском) распределении яиц хозяина и возможности многократного заражения при повторных встречах, доля незараженных хозяев равна:
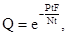 (10)
(10)
где е – основание натуральных логарифмов, P 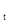 – плотность паразита в поколении t, N – плотность популяции хозяина в поколении t, F – плодовитость паразита.
– плотность паразита в поколении t, N – плотность популяции хозяина в поколении t, F – плодовитость паразита.
Соответственно, плотность популяции хозяина в поколении t+1 будет:
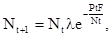 (11)
(11)
а плотность популяции паразита в поколении t+1 будет:
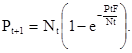 (12)
(12)
При такой ситуации возможны два выхода: либо продуктивность паразита окажется слишком мала, чтобы существенно снизить биотический потенциал хозяина – тогда обе популяции будут увеличивать свою численность, либо паразит уничтожит всех хозяев и вымрет сам. При введении в модель верхнего уровня численности хозяина, допускаемого ресурсами, модель предсказывает полное вымирание и хозяина, и паразита. Если ввести в формулу верхний порог численности паразита, вся система может существовать неограниченно долго. Однако модель Томпсона основана на неверной предпосылке о независимости количества яиц, отложенных паразитом, от плотности популяции хозяина. Этого недостатка лишена модель Л.Никольсона и В.Бейли (A.J.Nicholson, V.A.Bailey, 1935), которые также пытались объяснить имеющее обычно место в природе длительное сосуществование паразита и хозяина.
Эти авторы исходили из следующих предположений. В течение своей жизни самка паразита в поисках хозяина обследует определенную площадь – а. Величину этой площади можно считать в среднем постоянной и характерной для вида. Можно предположить далее, что поиск беспорядочен и траектория движения самки напоминает броуновское движение. Действительно, например, у трихограммы перемещение самок в поисках яиц хозяина происходит по сложной запутанной линии (рис.32) (отметим, однако, что при попадании на участок поверхности, загрязненной кайромоном – привлекательным для паразитов веществом, оставляемым бабочкой на поверхности при откладке яиц, характеристики движения трихограммы меняются и резко возрастает число поворотов). Другое предположение этих авторов – количество яиц, которое может отложить самка паразита, не ограничено по сравнению с числом хозяина.
Приняв эти предположения, перейдем к самой модели. Очевидно, что если плотность популяции хозяина N, а самка паразита обследует площадь а, то она встретит и соответственно заразит а N хозяев, т.е. ее плодовитость
F = a N.

Рис.32. Пример траектории движения трихограммы на горизонтальной плоскости. Точки нахождения трихограммы через определенные интервалы времени соединены прямыми (по В.Б.Чернышеву и др., 1988)
Подставив это значение в формулу Томпсона (12), получим, что число погибших от паразита хозяев, равное числу паразитов в следующем (поколении, будет:
 (13)
(13)
Если территория, обследуемая одной самкой, равна а, то группа самок P обследует площадь, меньшую произведения aP, так как траектории самок перекрещиваются друг с другом. Анализ этих случайных перекрытий обследованных территорий показывает, что имеет место следующее соотношение:
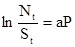 , (14)
, (14)
где N – исходное число хозяев, S – число хозяев, оставшихся незараженными. Соответственно, зная плотности популяций паразита и хозяина, а также процент зараженных хозяев, можно определить величину территории а, обследуемой одной самкой. Если каждая выжившая особь хозяина производит в среднем F 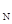 потомков (плодовитость хозяина), то плотность популяции хозяина в поколении t+1 будет равна:
потомков (плодовитость хозяина), то плотность популяции хозяина в поколении t+1 будет равна:
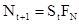 .
.
Эту формулу можно переписать следующим образом:
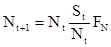
Логарифмируя это уравнение, получаем:
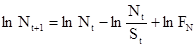 .
.
или, используя равенство (14):
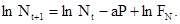 (15)
(15)
Пользуясь последней формулой, можно рассчитать плотность популяции хозяина в последующем поколении. Очевидно, что число паразитов в следующем поколении должно быть равно числу зараженных хозяев в предыдущем поколении.
Эта модель была проверена в теплице, где имелись белокрылки Trialeurodes vaporariorum и ее паразит Encarsia formosa (T.Burnett, 1958). Результаты этого эксперимента представлены на рис. 33. Можно видеть неплохое совпадение теоретически вычисленных и экспериментальных данных. Для подобных взаимодействий типично некоторое отставание во времени изменений популяции паразита, за счет чего и возникает колебательный процесс.
В принципе, вся система будет равновесной, если 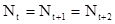 и т.д. Если подставить это равенство в формулу (15) мы получаем, что:
и т.д. Если подставить это равенство в формулу (15) мы получаем, что:
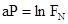

Рис.33. Взаимодействия между популяциями тепличной белокрылки Trialeurodes и ее паразита Encarsia в опытной теплице. Толстые линии – результаты наблюдений, тонкие линии – расчет по формуле (по Т.Бернету из Дж. Варли и др., 1978)
Однако эта стабильность
Дата добавления: 2015-05-16; просмотров: 1782;