Проба с мехолилом как показатель реактивности симпатической системы у человека 5 страница
ГЛАВА ft
или через интерорецепторы, чувствительные к подобным сдвигам. У животного появляется «насторожен ность» и возникает исследовательская двигательная активность [181].
Стеллар считает, что активность возбуждающих центров гипоталамуса регулируется многими факторами, которые можно объединить в 4 категории: 1) тормозные центры гипоталамуса, которые служат только для подавления возбуждающих центров; 2) сенсорные раздражители и афферентные импульсы, которые они вызывают; 3) гуморальные факторы внутренней среды, влияющие на гипоталамус че|рез сосудистую систему, и
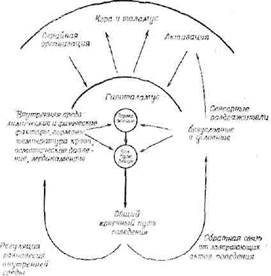
физиологических факторов в мотивации Объяснение см.
ПСИХИЧЕСКИЕ ПРОЦЕССЫ С ФИЗИОЛОГИЧЕСКОЙ ТОЧКИ ЗРЕНИЯ 247
4) центры коры и таламуса, которые могут оказывать тормозное и возбуждающее влияние на гипоталамус. Стеллар изобразил свои представления графически в простой .и ясной форме (фиг. 32). В центре схемы слева стоят физические и химические факторы внутренней среды — температура, гормональное равновесие, осмотическое давление и т. д., влияющие как на тормозные, так и па возбуждающие центры гипоталамуса. Активность центров любого типа может быть повышена или понижена такими физико-химическими влияниями. Следует отметить также, что эти факторы, которые имеют первостепенное значение для гипоталамических центров «мотивации поведения», являются одновременно продуктами поведенческих реакций. Справа показаны сенсорные раздражители, оказывающие прямое влияние на гипоталамус через афферентные пути. Природа их влияния зависит, однако, от определенных временных и пространственных характеристик и от изменений, вносимых предшествующими раздражителями (обучение). Сенсорные раздражения представляют собой комбинацию внешнего раздражения и раздражений, возникающих в результате участия различных органов в поведенческих реакциях. Кроме того, влияния коры и таламуса, прямо или косвенно возникающие при сенсорных раздражениях, а также от восходящих влияний самого гипоталамуса, действуют на активность гипоталамуса через различные соединительные пути. Понятно, что физиологическая регуляция гипоталамуса весьма сложна.
Специфические „системы поощрения"
Олдс [690] предпринял попытку установить, не делится ли обширная система положительной мотивации в мозге крысы на субсистемы более специфического характера, такие, как «система пищевого поощрения», «система полового поощрения» и т. д. Одна из его рабочих гипотез сводилась к следующему: «Если электрическое раздражение некоторых областей вызывает разряды клеток, опосредующих пищевое подкрепление, следует ожидать, что стремление животного к самораздражению будет снижаться и повышаться параллельно чувст-
ГЛАВА IX
ву голода, т. е. как аппетит». Первые исследования показали, что если электроды помещали в определенные точки вептро-медиального ядра гипоталамуса и в область перегородки, то голод повышал частоту самораздражений, т. е. выдвинутая Олдсом гипотеза подтверждалась.
В более поздней работе Олдс [690] применил метод, позволивший ему еще более строго локализовать систему пищевого поощрения. Модификация методики состояла в том, что вместо постоянной интенсивности раздражения были использованы раздражения разной силы: 0, 10, 15, 20, 25, 30, 35 и иногда 40 мка, каждая из которых в опыте применялась в течение 8 мин. У каждой крысы определяли число реакций за 8 мин в течение трех экспериментов, во время которых крыса была накормлена, и в течение трех экспериментов после 24-часового голодания. Оказалось, что, применяя раздражения разной интенсивности, мы получили разные карты «зон пищевого подкрепления». У одной из крыс с электродом в заднем отделе гипоталамуса частота самораздражений после 24-часового голодания оказалась на 200% выше, но подобная ситуация имела место лишь при определенной интенсивности раздражения — 10 мка. При раздражениях любой другой силы состояние насыщения не оказывало никакого влияния на частоту самораздражений. При другой локализации электрода частота само-раздражений в голодном и сытом состоянии была одинакова, если сила раздражений не превышала 20 мка. При всех более сильных раздражителях (25—40 мка) наблюдалось резкое расхождение в частоте самораздражений— на фоне голода она значительно возрастала. Составление карт с применением раздражений различной силы показало, что в заднем отделе гипоталамуса существует специфический «центр пищевого поощрения».
Олдс показал также существование «специфической системы полового поощрения». При вживлении электродов в дорсо-медиальную часть хвостатого ядра и достаточно высоком содержании андрогенов в организме наблюдается выраженная реакция (самораздражение) на ток 15 мка. После кастрации, когда уровень андрогенов
ПСИХИЧЕСКИЕ ПРОЦЕССЫ С ФИЗИОЛОГИЧЕСКОЙ ТОЧКИ ЗРЕНИЯ 249
снижается, частота самораздражепий также уменьшается. При очень низком уровне андрогенов животное совсем не реагировало даже при силе тока, доходящей до 55 мка. Затем вводилась большая доза тестостерона, и на следующий день животное реагировало на ток 55 мка, затем на 45 и, наконец, снова на 15 мка. Уровень андрогенов определяет наличие или отсутствие реакций, а также порог действия раздражителя, но когда реакция имеет место, частота самораздражений не зависит от силы раздражителя.
Другой эксперимент, проведенный в лаборатории Олд-са, выявил поразительные различия между специфическими системами подкрепления. У 16 животных измеряли влияние голода и уровня андрогенов на частоту самораздражений при токе 1 в (приблизительно 60— 80 мка) в течение 1 час. В перегородке, гипоталамусе и хвостатом ядре были найдены точки, для которых выявлены обратные отношения между влиянием голода и андрогенов. Если голод повышал частоту самораздра-жений, то андроген снижал ее, и наоборот.
Эти данные ясно показывают, что «система положительной мотивации» действительно состоит из ряда специфических «субсистем», таких, как система пищевого подкрепления, система полового подкрепления и т. д. Необходимо, однако, одно предостережение. Наряду с убедительными доказательствами в пользу специфичности существуют также данные, свидетельствующие о том, что специфическая система не представляет собой изолированного центра. Например, кроме центра пищевого подкрепления в заднем отделе гипоталамуса, точки «пищевого подкрепления» имеются в других частях мозга, расположенных более рострально.
В целом на основании имеющихся данных можно сделать вывод, что физиологические механизмы мотивации, очевидно, состоят из общего влечения, модулируемого специфическими влечениями. Общее влечение, очевидно, является следствием гуморальных и нервных влияний на состояние центральной нервной системы, особенно на состояние ретикуло-гипоталамической активирующей системы. Нервные влияния на эту систему возникают в рецепторах всего организма — в интеро-
ГЛАВА IX
экстеро- « проприорецепторах, а также по механизму «обратной связи» от корковых и подкорковых центров. Специфические влечения, вероятно, являются следствием нервных и гуморальных влияний на специфические системы двух типов: 1) систему поощрения и 2) систему наказания. Наивысшая степень специфичности систем поощрения и наказания наблюдается в гипоталамусе и лимбической системе. Хотя is этом обсуждении мы почти полностью игнорировали теорию обучения, следует упомянуть о теории «снижения влечения», выдвинутой Халлом. Согласно этой теории, «подкрепление», способствующее обучению, зависит от снижения влечения, которое включает и регулирует поведение. Это объяснение, очевидно, приложимо только к системе наказания, которой, как теперь выяснилось, отведено в мозге сравнительно мало места. Сложнее решить, каким образом подавляется более обширная система поощрения, которая, как было показано, является мощным «возбудителем» поведения и обусловливает положительную обратную связь. Сама по себе положительная обратная связь приводит к постоянному повышению активности и в конце концов к разрушению механизма. Поэтому должна действовать также «отрицательная обратная связь». Несомненно, что удовлетворяющие влечение акты поведения играют роль в возникновении такой отрицательной обратной связи посредством гуморальных и нервных сдвигов, но подробности этих влияний неизвестны. Удовлетворение, если его определить операционально, представляет собой ослабление мотивированного поведения в результате достижения цели. Это в корне отличается от снижения интенсивности реакций, связанных с мотивированным поведением, вследствие активации «системы наказания». Возможно, что в обоих случаях подавляется активность механизмов положительной мотивации, но следует учитывать вероятность того, что положительная, мотивация может сохраняться, но маскироваться более сильной «отрицательной мотивацией».
В последнее время появился новый экспериментальный подход к проблеме мотивации— регистрация изменений активности различных отделов систем положительной я отрицательной мотивации под ;влиянием раз-
ИЧЕСКИЕ ПРОЦЕССЫ С ФИЗИОЛОГИЧЕСКОЙ ТОЧКИ ЗРЕНИЯ 251
дражителей, вызывающих или подавляющих мотивированное поведение. Портер и др. [721] обнаружили изменения электрической активности передне-латерального гипоталамуса при раздражении влагалища у кошки в период течки. Сойер и Кавакамп [525, 792а] зарегистрировали у самки кролика сдвиги электрической активности различных областей коры и подкорки (в том числе новой и лимбической коры, гиппокампа, перегородки, гипоталамуса и ретикулярной формации) при искусственном раздражении влагалища, а также во время и после покрытия. Особый интерес представляют открытые ими тонкие изменения внешнего поведения, которые были замечены только после того, как изменения в ЭЭГ заставили провести тщательные наблюдения за их поведением. Необходима большая экспериментальная работа в этом направлении.
ПОЗНАВАТЕЛЬНЫЕ ФУНКЦИИ
Со времен Платона писатели и философы выделяли по крайней мере два элемента, из которых складывается психика человека,— чувства и рассудок. В поведении человека проявляются оба эти элемента; рациональное и иррациональное поведение характерно для каждого из нас, количественно различаясь у разных индивидуумов, а также у одного и того же лица в различных условиях. Выше мы пытались дать очерк физиологии эмоций, показать важное значение подкорковых структур, особенно гипоталамуса, и' относительно недифференцированных древних корковых структур, таких, как обонятельный мозг. В организации эмоциональных реакций прежде всего участвуют ощущения, отражающие состояние организма. Боль вызывает эмоциональные реакции; прикосновение к коже и ее поглаживание вызывает успокоение у ребенка, даже лишенного больших полушарий, а вкусовые раздражения тесно связаны с приятными и неприятными ощущениями. Наоборот, восприятие через дистантные рецепторы — органы зрения и слуха — относится к окружающему нас миру .и формирует важнейшие элементы для интеллектуальных операций и для процесса обучения в целом. Вероятно, их
ГЛАВА IX
эмоциональное влияние незначительно, так как эти ощущения дают информацию не о .нашем организме, а об окружающей нас среде. Если раздражение тех органов чувств, которые вызывают эмоциональное возбуждение, приводит к немедленному рефлекторному действию, то при действии высших органов чувств — глаза и уха — этого не происходит, и именно эта задержка, отсутствие реакции, 'Непосредственно связанной с определенным раздражителем, и составляет основу разумного поведения '.
Мы не ставили своей задачей рассмотрение разумного поведения с точки зрения физиологии (откровенно говоря, мы не считаем себя способными выполнить эту задачу). Мы ограничимся лишь некоторыми замечаниями относительно восприятия и обучения с целью показать .роль больших полушарий (новая кора) и продемонстрировать неразрывную связь эмоционального и интеллектуального поведения. Поскольку основная цель данной книги — изучение эмоций, то мы рассмотрим далее роль гипоталамуса в различных кортикальных процессах.
Физиология восприятия
СПЕЦИФИЧЕСКИЕ СЕНСОРНЫЕ ПУТИ И КОРКОВЫЕ ПРОЕКЦИОННЫЕ ЗОНЫ
Ранее было упомянуто, что афферентные системы достигают коры головного мозга и заканчиваются в так называемых проекционных зонах, которые специфичны для каждой сенсорной «модальности». Так, раздражение сетчатки светом или раздражение сетчатки и зрительного нерва (а также любой части афферентного
' Чувство обоняния в этом отношении стоит особняком среди дистантных .рецепторов. Оно связано с древним (обонятельным) мозгом, а не с новой корой больших полушарий; оно играет важную роль в инстинктивных реакциях животных, но не имеет большого значения для человека. Решающую роль в таком развитии сыграл, по-видимому, переход к прямохождению, при котором обоняние утратило свое прежнее значение. Это нашло свое отражение и в малых размерах обонятельного мозга по сравнению с общим объемом мозга человека.
ПСИХИЧЕСКИЕ ПРОЦЕССЫ С ФИЗИОЛОГИЧЕСКОЙ ТОЧКИ ЗРЕНИЯ 253
пути, соединяющего сетчатку с проекционной зоной коры) электрическим током приводит № возбуждению части затылочной коры. Соответствующее возбуждение слухового аппарата вызывает активацию височной коры. Эти проекционные зоны различных сенсорных систем были определены в тончайших деталях при записи электрических потенциалов, возникающих в коре в ответ на зрительные, слуховые и другие сенсорные раздражения. Спонтанная электрическая активность мозга бодрствующих и наркотизированных животных маскирует «вызванные» потенциалы, возникающие при действии сенсорных раздражителей. Ряд авторов разработал простую методику, позволяющую обойти эти трудности. У животных вызывают глубокий наркоз, при котором спонтанная активность коры практически исчезает. При этих условиях специфические проекционные зоны коры продолжают получать импульсы от рецепторов и явля-ются удивительно чувствительными индикаторами импульсов, приходящих по специфическим афферентным путям. Например, такое слабое раздражение, как прикосновение к одному волоску на коже руки, может вызвать отчетливый потенциал в коре. Эти наблюдения ясно показывают, что активация проекционных зон коры сенсорными раздражителями недостаточна для восприятия, поскольку такая активация сохраняется в состоянии очень глубокого наркоза, когда сознание выключено. В этих условиях, а также во время глубокого сна ретикулярная формация не отвечает на звуковые щелчки [138, 498], тогда как у бодрствующего животного сенсорные раздражители вызывают реакцию как в сенсорной проекционной зоне коры, так и в ретикулярной формации.
Эту работу дополнили опыты с удалением специфических афферентных систем , хирургическим путем. Перерезка ножек нижнего двухолмия хотя и лишает слуховую кору специфических импульсов, идущих через лем-нисковую систему, но совершенно не изменяет реакций коры на звуковые раздражители [277]. Этот удивительный результат показывает, что, насколько можно судить по электрической активности, специфические классические слуховые пути и неспецифическая диффузная
254 ГЛАВА IX
ретикулярная формация имеют одинаковое значение. Следует напомнить, что при повреждении ретикулярной формации и латеральных частей среднего мозга (перерыв специфических афферентных путей) соответственно реакция пробуждения в ЭЭГ сохранялась у обоих видов препаратов, хотя животные с повреждением ретикулярной формации находились ,в состоянии комы, а те, у которых были повреждены специфические системы, казались нормальными. Однако надлежащая оценка сенсорных раздражений требует активности коры, которая не обнаруживается при регистрации реакции пробуждения в ЭЭГ или изучении корковых вызванных потенциалов. Это объясняет, почему при исследовании поведения животных, лишенных специфических сенсорных путей, обнаруживались серьезные дефекты сенсорного восприятия; они были «неспособны ко всему разнообразию форм поведения, включающему поддержание внимания, а также аффективные и приспособительные реакции» [857]. Эти наблюдения позволяют предположить, что восприятие, т. е. оценка сенсорного раздражения, выявляющаяся в соответствующем действии, требует какого-то взаимодействия между специфической и диффузной афферентными системами.
АКТИВИРУЮЩАЯ РЕТИКУЛЯРНАЯ СИСТЕМА И ВОСПРИЯТИЕ
Можно предположить, что неспецифическая (ретикулярная) афферентная система возбуждает кору и позволяет ей использовать сигналы, получаемые проекционными специфическими зонами по специфическим сенсорным путям, и воздействовать на эти сигналы. Согласно этому предположению, импульсы, приходящие по неспецифической системе, подготавливают кору к осознанию чего-то; однако без импульсов, идущих по специфической системе, ей нечего осознавать. Очевидно, при приходе афферентных импульсов только по специфической или только по неспецифической системе ничто не воспринимается.
Есть, однако, все основания задать вопрос: действительно ли ретикулярная формация является той неспецифической афферентной системой, которая имеет
ПСИХИЧЕСКИЕ ПРОЦЕССЫ С ФИЗИОЛОГИЧЕСКОЙ ТОЧКИ ЗРЕНИЯ 255
такое значение для процесса «восприятия? У человека существует прямая связь между интенсивностью светового раздражения, амплитудой вызванного потенциала зрительной коры и интенсивностью зрительного восприятия [444]. Это позволяет считать, что взаимодействующие системы, участвующие в восприятии, облегчают корковые реакции на сигналы, действующие через специфические проекционные системы. Однако многие авторы показали, что раздражение ретикулярной формации снижает потенциалы, вызываемые в специфической проекционной зоне коры [126, 766]. В соседних ассоциативных зонах этот эффект еще более выражен [126]. Поэтому кажется вероятным, что взаимодействие между ретикулярными неспецифическими афферентными импульсами и импульсами специфических проекционных путей служит какой-то другой функции, а не восприятию. Предполагается, что это взаимодействие способствует концентрации внимания путем снижения реактивности коры в зонах, не связанных с тем определенным восприятием или действием, которое выполняет организм в данный момент. Вероятно, это достигается с помощью торможения где-то на афферентном пути, что приводит к снижению афферентного притока к коре, i Можно сказать, что, хотя восприятие без реакции активации невозможно, очевидно, не реакция активации, возникающая при действии активирующей ретикулярной системы, обеспечивает механизм, необходимый для ' восприятия. Она способствует восприятию путем снижения посторонних влияний, но источник коркового облегчения, которое при взаимодействии с приходящей специфической сенсорной информацией создает восприятие, мы должны искать где-то в другом месте.
ВОСПРИЯТИЕ И ГИПОТАЛАМУС
Как будет показано ниже, эмоции и восприятие тесно связаны между собой. В свете этого факта можно предположить, что в их основе лежит какой-то общий нервный процесс. Возможно, что возбуждение гипоталамуса взаимодействует с сенсорным притоком к коре и повышает корковые реакции на импульсы, достигающие ее
25b
ГЛАВА IX
по специфическим проекционным путям. Следующие наблюдения подтверждают эту гипотезу.
В опытах на животных реакция специфических проекционных зон коры на световые и звуковые раздражения облегчается при одновременном раздражении заднего отдела гипоталамуса. Слабые сенсорные раздражения, которые в контрольных опытах оказываются неэффективными в 50% проб или более, регулярно вызывают корковый ответ при их нанесении на фоне активации гипоталамуса. Частота стрихиинных разрядов, вызываемых локальным нанесением этого вещества на зрительную или слуховую кору, повышается при сочетании раздражения гипоталамуса со специфическими сенсорными раздражениями. Этот эффект сохраняется в течение некоторого времени после прекращения раздражения; его нельзя приписать сдвигам кровообращения, поскольку он имеет место даже в отсутствие изменений кровяного давления [347].
Аналогичный результат был получен при рефлекторном возбуждении гипоталамуса болевыми раздражителями [349]. Даже подпороговые раздражения, которые не влияют на корковые потенциалы, резко повышают интенсивность реакции специфических проекционных зон коры на звуковые (фиг. 33) или зрительные раздражения. Чрезвычайно интересно, что под влиянием гипота-ламического или болевого раздражения сенсорная реакция возникает в таких областях проекционной зоны, в которых она при контрольном опыте отсутствует. Отсутствие такой суммации при глубоком барбитуратном наркозе зависит от чувствительности мультисинаптиче-ской гипоталамической системы к барбитуратам [304].
Это наблюдение удовлетворяет некоторым требованиям физиологической теории процесса восприятия, поскольку: 1) реактивность сенсорной проекционной зоны повышается под влиянием прямой или рефлекторной активации гипоталамо-кортикальной системы; 2) наркоз, влияющий на эту активацию, устраняет подобную сум-мацию. Кроме того, на кошках с электродами в заднем отделе гипоталамуса можно показать, что раздражение этой области слабым электрическим током, вызывающее только расширение зрачка и сокращение третьего
ПСИХИЧЕСКИЕ ПРОЦЕССЫ С ФИЗИОЛОГИЧЕСКОЙ ТОЧКИ ЗРЕНИЯ 257
века, чрезвычайно повышает реакцию на сенсорные раздражения. Звуковые или световые сигналы, подаваемые в различных местах вблизи от кошки, вызывают поворот головы по направлению к ним во время раздражения гипоталамуса, но не оказывают действия сразу же после прекращения этого раздражения [310]. Очевидно, восприятие является следствием взаимодействия между
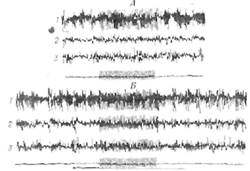
Фиг. 33. Изменение реакции коры на звуковое раздражение пол действием боли (347].
Л. Звуковое раздражение (щелчок) в контрольном опыте. В. Звуковое раздражение при погружении правой задней лапы в воду при 55° (погружение отмечено длинной горизонтальной линией). Калибровка: 300 мкв и 1 се/с. / и 2 — левая эктосильвиева извилина: 3— правая эктосильвиева извилина.
возбуждением коры под влиянием сенсорного раздражения и гипоталамо-кортикальным разрядом, возникающим при прямом раздражении гипоталамуса или при рефлекторной активации гипоталамической системы. Кроме того, эти наблюдения показывают, что эмоциональное возбуждение может участвовать в процессе восприятия и, вероятно, изменять его.
Прежде чем перейти к рассмотрению некоторых пси-хо-физиологических наблюдений, подтверждающих наше мнение об участии гипоталамо-кортикальной системы
9-517
ГЛАВА IX
в восприятии, необходимо подробнее обсудить роль ретикулярной формации.
Мы неоднократно подчеркивали, что ретикулярная формация и гипоталамус тесно связаны между собой анатомически и функционально [243, 680]. Многие специфические функции, обслуживаемые гипоталамусом, делают необходимым разграничение этих двух структур, но, учитывая многочисленные пути, связывающие их, можно ожидать, что в интактном организме возбуждение ретикулярной формации должно сопровождаться возбуждением заднего отдела гипоталамуса. Вполне возможно поэтому, что облегчающее действие гипоталамуса на сенсорные проекционные зоны должно проявляться и при раздражении ретикулярной формации. Эксперименты действительно показали, что при раздражении ретикулярной формации в одних нейронах зрительной зоны коры отмечается эффект облегчения, а в других — эффект торможения. Джаспер и Ажмон-Мар-сан [514], а также другие авторы [160, 572] обнаружили, что такое облегчение возникает при активации ретикулярной системы на уровне таламуса, а не на уровне среднего мозга, а Бремер и Ступель [102] отметили, что эффект облегчения может быть вызван через ретикулярную формацию, если специфическая сенсорная система активируется раздражением таламических ядер переключения, тогда как при раздражении рецепторов возникает торможение.
Хотя мы еще не можем перечислить все факторы, котчрые ответственны за облегчение и торможение, весьма возможно, что эффект облегчения может быть вызван с ретикулярной формации на всех уровнях, но при раздражении ее на уровне среднего мозга нисходящий разряд тормозит переключающие ядра. Возможно, именно по этой причине эффект облегчения легче вызвать с таламуса и гипоталамуса. Проведенные недавно исследования показали, что снижение сенсорного притока может быть достигнуто также и с промежуточного мозга [189], но, очевидно, участвующие в этом области не перекрываются с теми, с которых вызывается диффузное возбуждение коры и облегчающее действие на сенсорные проекционные зоны.
ПСИХИЧЕСКИЕ ПРОЦЕССЫ С ФИЗИОЛОГИЧЕСКОЙ ТОЧКИ ЗРЕНИЯ 259
В литературе отмечалось, чт*о для восприятия необходима не только общая активация и что особое значение имеют процессы возбуждения и торможения. Так, Л инд слей [581] различает общую реакцию пробуждения (general alerting — переход от сна к бодрствованию) и специфическую активацию (specific alerting), которая связана с вниманием и восприятием. В последнем случае одни области коры подвергаются торможению, а другие — возбуждению. Мы считаем, что ретикулярная формация среднего мозга вызывает реакцию пробуждения, а также участвует в снижении сенсорного притока к коре, что делает возможным концентрацию внимания на некоторых объектах; последний процесс включает, кроме того, облегчающее влияние гипоталамуса (и та-ламуса?) на некоторые сенсорные проекционные зоны. Тот факт, что в условиях возбуждения гипоталамуса сенсорные раздражители активируют значительно более обширные зоны коры, чем в отсутствие активации гипо-таламо-кортикальной системы, может быть основой процесса связывания объекта, действующего на сенсорный орган с аналогичным предшествующим сенсорным опытом (узнавание) '.
Ранее мы упоминали, что под влиянием раздражения гипоталамуса подпороговые раздражители вызывают поведенческие реакции. Из этого наблюдения следует вывод, что эмоциональные реакции, явные или скрытые, играют важную роль в восприятии, и эта точка зрения подтверждается различными экспериментами.
* * * *
' Хотя мы подчеркнули значение активации гипоталамуса для возникновения облегчения корковых сенсорных потенциалов, которое кажется нам основой восприятия, форма (и степень) сенсорного возбуждения, очевидно, также имеет существенное значение. Этим можно объяснить тот факт, что раздражение ретикулярной формации на фоне активации зрительной коры мелькающим светом приводит к торможению при низкой частоте световых мельканий, но к облегчению при высокой их частоте [868]. Кроме того, раздражение ретикулярной формации, снижающее зрительные корковые потенциалы в контрольных условиях, может повышать их после значительного снижения эффективности зрительного раздражителя под влиянием привыкания [623].
9»
260 ГЛАВА IX
В опытах по выработке условных рефлексов звук предшествовал слабому электрическому раздражению, наносимому через решетку, на которой стояла кошка. Это сочетание многократно повторялось, что приводило к предвосхищению болевого ощущения при применении звука. Эта условнорефлекториая реакция проявлялась характерным изменением ЭЭГ и эмоциональной реакцией (ворчание) при восприятии звука. Однако, если звук повторялся много раз без «подкрепления» электрическим ударом, эмоциональная реакция и корковый ответ, наблюдаемый по ЭЭГ, исчезали [278]. Таким образом, судя по этому примеру, восприятие связано с эмоциональной реакцией, которая, как мы говорили ранее, включает гипоталамическую реакцию и гипоталамо-кортикальное взаимодействие.
Фустер [274] выработал у обезьян диффереицировку между двумя зрительными объектами — причем правильный выбор подкреплялся пищей,— а затем постепенно уменьшал время экспозиции. Он обнаружил, что раздражение ретикулярной формации снижает время реакции и повышает число правильных реакций. Вряд ли можно сомневаться, что в этом эксперименте, как в любом другом опыте с условными рефлексами, где подкреплением служит пища или страх перед наказанием, ги-поталамическая система будет адекватно реагировать на раздражение ретикулярной формации. Повышение скорости реакции и улучшение зрительного различения объектов при краткой экспозиции вполне могут быть выражением облегчающего действия гипоталамуса на сенсорную кору, которое в этом эксперименте первично вызывается раздражением ретикулярной формации.
В качестве дальнейшего подтверждения нашего тезиса о том, что гипоталамо-кортикальные разряды участвуют в восприятии, можно упомянуть о некоторых наблюдениях, сделанных на людях. У больного, находящегося в состоянии комы, в ответ на бессмысленный слуховой раздражитель реакции в ЭЭГ не возникает, однако, если произнести имя больного, такая реакция появляется [560]. Значение собственного имени — биологическое и эмоциональное, — очевидно, достаточно
•ИХИЧЕСКИЕ ПРОЦЕССЫ С ФИЗИОЛОГИЧЕСКОЙ ТОЧКИ ЗРЕНИЯ 261
вепико, чтобы вызвать реакцию даже в глубоко заторможенной коре. Другой иллюстрацией нашего тезиса, сог-1асно которому эмоциональное состояние (говоря языком физиологии, гипоталамо-кортикальный разряд) определяет реактивность мозга к сенсорным раздражителям, может служить общеизвестный факт: мать просыпается при малейшем шорохе, доносящемся из кроватки ребенка, и не реагирует на значительно более сильные, но эмоционально безразличные звуковые раздражители.
Наконец, психологи показали, что хотя слова, предъявляемые испытуемым визуально на долю секунды, не могут быть узнаны, те из них, которые обладают эмоциональным содержанием, вызывают потоотделение на ладонях [615]. Это кожногальванический рефлекс, обычно наблюдаемый при эмоциях. Этот и другие подобные эксперименты подтверждают справедливость положения, что «аффективные реакции предшествуют рациональным» [646], и иллюстрируют значение эмоциональной реактивности для' восприятия. Мы считаем возможным завершить обсуждение физиологических основ восприятия следующими общими замечаниями.
Дата добавления: 2014-12-10; просмотров: 1132;