Проба с мехолилом как показатель реактивности симпатической системы у человека 4 страница
МОТИВАЦИЯ
В самом широком смысле мотивация включает все факторы, которые вызывают и направляют акты поведения '. Однако здесь мы рассмотрим не факторы среды, воздействующие на организм, а внутренние состояния и процессы. Вопрос заключается в том, каким образом возникают изменения внутренних состояний и процессов и как они влияют на поведение.
Итак, прежде всего перед нами задача дать определение поведения. Где следует провести границу между внешне наблюдаемыми формами активности, которые, несомненно, относятся к категории поведения, и «внутренними состояниями и процессами», которые вызывают и направляют его? Единственное возможное место для такой условной границы, очевидно, лежит между теми видами активности, которые можно наблюдать по внешним проявлениям, и теми, которые наблюдать нельзя. Таким образом, мы будем считать вместе с Хеббом [437], что поведение состоит из внешне наблюдаемых форм активности организма. Понять механизмы поведения можно лишь путем изучения внутренних сдвигов, а для этого необходимы самые разнообразные методы и приборы. Поведение в том смысле, в котором употребляем это слово мы, включает как рефлекторные, так и произвольные акты, но не охватывает изменений внутренней среды или центральной нервной системы, которые предшествуют актам поведения и вызывают их.
Термин влечение был первоначально использован, для обозначения тех внутренних сдвигов, с которыми * * * *
1 В этом смысле мотивация имеет то же значение, что и влечение у Моргана [662].
ПСИХИЧЕСКИЕ ПРОЦЕССЫ С ФИЗИОЛОГИЧЕСКОЙ ТОЧКИ ЗРЕНИЯ 231
связаны биологические потребности, такие, как голод, жажда и половое влечение, и которые вызывают поведение направленное на удовлетворение этих потребностей. По мере расширения биологических исследований перечень биологических потребностей значительно увеличился и пришлось приложить большие усилия, чтобы показать, что все виды мотивации целиком исходят из очень немногих действительно основных влечений, таких, как голод, жажда и половое влечение.
В дальнейшем были предприняты попытки создать теорию «единого влечения». Одна из таких теорий считает, что все примитивные мотивации можно свести к стремлению избавиться от некоторых форм болевого воздействия [437]. Чувство голода рассматривали как стремление избавиться от «голодных колик» — боли, возникающей при сокращениях пустого желудка. Инстинкт материнства связывали с возникновением «болей» при растяжении молочной железы, жажду — с сухостью во рту и глотке и т. д. Однако сохранение материнского инстинкта после удаления молочных желез, чувства голода после удаления желудка и жажды без сухости во 'рту и глотке, а также многие другие данные опровергают эту теорию. Все виды мотивации имеют общие черты, и ниже мы приведем соответствующую теорию .мотивации, основанную на изученных физиологических механизмах.
Очевидно, в основе всех видов мотивации независимо от того, участвуют в ней внешние раздражения или нет, и независимо от направления вызываемого ими поведения лежит активация мозга. Кроме этой генерализованной активации, по-видимому, существуют специфические механизмы, которые направляют поведение и в определенных условиях вызывают специфические виды поведения. Хебб [437] говорит об «актива-ционной функции» и «ключевой функции» механизмов мотивации и возбуждающих их раздражителей. Таким образом, проблема сводится к выявлению физиологических механизмов этих двух аспектов мотивации. Несомненно, что к этим механизмам относятся гуморальные сдвиги, но мы можем считать, что такие сдвиги воздействуют на центральную нервную систему, которая регу-
ГЛАВА IX
лирует поведение. В связи с этим мы будем рассматривать поведение организма в целом, а не те фрагменты поведения, какими являются рефлекторные акты, осуществляемые через относительно короткие и прямые рефлекторные дуги. Иными словами, мы оставляем безусловные рефлексы за пределами нашего анализа. Наоборот, условные рефлексы, несомненно, входят в состав мотивированного поведения, так как очевидно, что выработка условных рефлексов изменяет внутреннее состояние, которое определяет .поведение. Не имеет значения, «произвольно» ли организм избирает ту или иную форму поведения, или «выбор» осуществляется автоматически на основе ранее выработанной «программы». Не имеет значения (для нашего рассуждения) и то, является ли эта «программа» врожденной или приобретенной — существует много данных, свидетельствующих о ее двоякой .природе. Весьма важно ответить на вопрос о том, каким образом и в каких пределах поведение организма вызывается и направляется условиями и процессами в центральной нервной системе, а не является просто комплексом вызываемых определенными сигналами рефлекторных актов, детерминируемых исключительно совокупностью внешних раздражений. Простые рефлекторные акты относительно легко понять, поскольку в их основе лежат более или менее прямые связи, между рецепторами и эффекторами с незначительным числом вставочных нейронов. .
В ситуациях, в которых возможен выбор между различными реакциями, анализ поведения не столь прост. Реакцию значительно труднее предвидеть, поскольку она определяется не только действующими в данное время 'раздражителями, но их взаимодействиями в центральной нервной системе со следами бесчисленных ранее действовавших раздражителей. Поведенческие реакции организма обусловлены прошлым опытом. Кроме того, между афферентными и эфферентными периферическими путями лежат сложные нейронные сети мозга. Нередко кажется, что какой-либо акт проявляется de novo в отсутствие явных «запускающих» его раздражителей. Часто можно отчетливо видеть, что на организм одновременно действует множество раздражителей.
!Е ПРОЦЕССЫ С ФИЗИОЛОГИЧЕСКОЙ ТОЧКИ ЗРЕНИЯ 233
В любом случае возникают два основных вопроса: п что заставляет животных реагировать тем или иным образом и 2) что заставляет их выбирать или составить особый вид поведения из имеющегося в их распоряжении набора .реакций.
Системы положительной мотивации
Исследования последнего времени показывают, что электрическое раздражение некоторых областей мозга может иметь для животного «мотивационное» значение. Олдс [690] обнаружил в опытах на крысах стремление к повторному раздражению области передней комиссуры через вживленные электроды; каждый раз, когда крыса случайно заходила в определенный угол ящика, она получала раздражение. После нескольких повторений крыса начинала упорно возвращаться в тот угол, где она получала явно желаемое раздражение.
В более тонкой модификации этих экспериментов был использован ящик Скиннера с рычагом; нажимая на рычаг, животное может произвольно раздражать свой мозг через хронически вживленные электроды. Первые эксперименты, проведенные этим методом, показали, что в мозге крысы, по-видимому, существуют обширные области, принадлежащие к «системе поощрения»; возбуждение этих областей вызывает стремление повторить действие раздражителя. Протяженность этой системы очень велика; некоторое время даже считали, что крыса стремится к самораздражению при помещении электродов в любую точку мозга.
Вскоре, однако, стало ясно, что система поощрения, выявленная в этих предварительных экспериментах, в общем соответствует проекциям системы свода, а при полном ее исследовании выяснилось, что она занимает практически весь обонятельный мозг вместе с гипоталамусом и связанными с ним структурами (фиг. 31). В сумме к этой системе принадлежит около 35% мозга крысы.
За годы, прошедшие после первого сообщения о роли электрического раздражения мозга как положительного подкрепления [691], аналогичное явление было про-
ГЛАВА IX
демонстрировано на других животных и обнаружено у человека. У кошек такое положительное подкрепление наблюдалось при раздражении через электроды, вживленные в хвостатое ядро [833]. Сходные результаты были получены с опытах на обезьянах с электродами в различных частях мозга, главным образом в лимбиче-ской системе [125]. Теперь хорошо известно, что животные многих видов охотно наносят самораздражение через электроды, вживленные в различные точки лимбиче-ской системы и связанных с нею структур [91].
У больных раздражение области перегородки сопровождается некоторым возбуждением и ускорением речи, причем, по их словам, они себя «очень хорошо чувству-
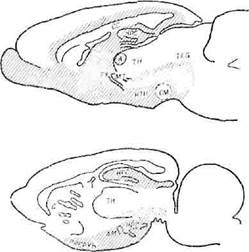
Фаг. 31. Области положительной (штриховка) л отрицательной (точки) мотивации у крысы, определяемые в опытах с самораздражением 1690].
ЕСКИЕ ПРОЦЕССЫ С ФИЗИОЛОГИЧЕСКОЙ ТОЧКИ ЗРЕНИЯ 235
ют» [435]. Особенно интересно, что при невыносимых болях раздражение такого рода сразу же вызывало облегчение, которое удавалось поддерживать путем повторных раздражений. В одном случае после 3-го раздражения наблюдалось стойкое улучшение, но этот случай представлял исключение. Возникает вопрос, каков механизм этого явления,— то ли боль просто маскируется «положительно подкрепляющим» эффектом удовольствия, то ли происходит блокада восходящих импульсов. По мнению самих больных, речь идет об истинном исчезновении болей, а не о появлении безразличия к болевым ощущениям, характерного для больных, перенесших лоботомию. В этих случаях сдвиг периферического болевого порога (например, при уколе булавкой) лишь незначителен или вообще отсутствует. Стеллар [864] в своем прекрасном обзоре «Физиология мотивации» высказывает предположение, что при избегании боли; решающую роль в мотивации, по-видимому, играет лобная кора. Это вполне разумное объяснение. Невозможно узнать, действительно ли раздражение области перегородки устраняет боль у животных, как это наблюдается у человека [435], или оно просто тормозит процессы, связанные с внешним выражением ощущения боли. Естественно, что этим ставится под сомнение та гипотеза, которую выдвинул Дельгадо [177], объяснивший внезапную потерю интереса к пище, обнаруживаемую у обезьян при раздражении перегородки, как результат торможения мотивации; ведь если у человека речь идет об устранении восприятия боли, то и результаты опытов Дельгадо можно объяснить как следствие подавления не мотивации, а восприятия.
В литературе было описано, что в мозге человека существуют «зоны удовольствия» (вероятно, тесно связанные с лимбической системой), раздражение которых вызывает различные виды приятных ощущений: спокойствие и облегчение, ощущение радости, ощущение глубокого удовлетворения [814]. Следует, однако, указать, что зоны наказания и поощрения лежат недалеко друг от Друга. При помещении электродов в «зону поощрения» больные доводили себя самораздражением до судорог, после которых они лежали расслабленные, со счастли-
236 ГЛАВА IX
вой улыбкой [814]. Раздражение височных долей вызывало выражение удовольствия, смех и шутливые замечания [177].
Интенсивность положительной мотивации, которая может быть достигнута путем раздражения «системы поощрения», удивительна. В экспериментах Олдса частота самораздражений у крыс с электродами, вживленными в область покрышки, достигала 7000 в 1 час, а при вживлении электродов в задний отдел гипоталамуса непосредственно кпереди от маммилярных тел — 5000 в 1 час. Чем дальше кпереди помещали электроды, тем ниже становилась частота самораздражения (в пределах 400—1100 в 1 час) [690].
Прямая связь между интенсивностью раздражения и частотой самораздражения была показана Олдсом для многих изученных им областей мозга. Он объясняет это вовлечением (активацией) нейронов системы поощрения в сферической зоне, радиус которой зависит от силы раздражителя. Он полагает, что отдельные нейроны подчиняются закону «все или ничего» и что вовлечение дополнительных нейронов является единственным фактором, благодаря которому возрастает мотивационное влияние при усилении интенсивности раздражителя. Однако у нас нет оснований сомневаться в том, что частота разрядов одиночных нейронов также изменяется при изменении силы раздражителя, поскольку это характерно для всех центральных нейронов. Вероятно, частота разрядов и пределы вовлечения вместе определяют интенсивность эффекта.
Раздражение «системы поощрения» электрическим током, по-видимому, создает более выраженную мотивацию, чем «естественное» поощрение (подкрепление). Стремясь к рычагу для «самораздражения», крыса «шла на то», чтобы подвергнуться болезненному удару электрического тока значительно большей силы, чем тот, который способна переносить голодная крыса, стремящаяся достичь пищи [690].
Описаны поразительные результаты раздражения «систем положительной мотивации» у обезьян [576]. При нанесении через вживленные электроды серии сдвоенных электрических импульсов были выявлены области
[ЧЕСКИЕ ПРОЦЕССЫ С ФИЗИОЛОГИЧЕСКОЙ ТОЧКИ ЗРЕНИЯ 237
мозга, раздражение которых воспроизводит некоторую заученную форму поведения; Лилли называет такую форму «старт-реакцией». Например, экспериментатор нажимает на ключ и наносит первую серию раздражений. У обезьяны возникает мотив к самораздражению, и «она становится частью циклической системы с обратной связью, из которой ее нельзя удалить до тех пор, пока возможно включение раздражителей». Если длительность серий импульсов, наносимых при однократном нажиме на ключ, снижается, то частота, с которой обезьяна нажимает на ключ, возрастает, т. е. животное стремится к максимальному возбуждению «системы поощрения». Одна обезьяна с электродами, «вероятно, находившимися в нижней части покрышки среднего мозга», доводила себя до полного изнеможения. В этом эксперименте каждая серия импульсов вызывала сокращение мышц во всем теле. «Обезьяна научилась использовать эти сокращения для механического замыкания ключа и довела частоту самораздражений до 18 в 1 сек в течение коротких периодов. После каждого такого периода активности ее рука падала с ключа, и она издавала звук «вопросительного» характера, после чего следовала пауза, и начинался новый период активности. Таким образом она «работала» до полного изнеможения и засыпала спустя 20 час, произведя в общей сложности около 200 000 серий импульсов». Следующее место по эффективности «положительной мотивации» занимало вентро-медиалыюе ядро гипоталамуса, а более слабые положительные реакции можно было получить (в порядке уменьшения эффективности) с интралами-нарной системы таламуса, перегородки, верхней части свода, головки хвостатого ядра и скорлупы. Исследование Лилли [576] показало, что исключительно активные области положительной мотивации находятся в гипоталамусе и среднем мозге.
Общие системы отрицательной мотивации
Под «системами отрицательной мотивации» мы подразумеваем те системы мозга, возбуждение которых действует как наказание и вызывает поведение, явно
ГЛАВА IX
направляемое желанием избежать их активаида. Как и «системы поощрения», эти «системы наказания», очевидно, преимущественно связаны с гипоталамо-лимбиче-скими структурами.
Сели области мозга, раздражение которых у крысы является «поощряющим», занимают значительную часть мозга (357о), то области, раздражение которых действует как наказание, составляют всего 5% мозга; животное делает отчетливые попытки избежать раздражения этих областей. Обозначенные точками области на фиг. 31 представляют ретикулярную формацию среднего мозга и части таламуса и гипоталамуса. Тенденция избегать электрического раздражения наблюдалась «только в некоторых участках этой очень ограниченной системы».1 Относительно небольшие размеры «системы наказания» по сравнению с обширной «системой поощрения» дают основания для важных ьыводов в теориях, касающихся обучения.
Дельгадо [177] показал, ч iо раздражение некоторых точек гиппокампа, задне-вентрального ядра таламуса, покрышки и центрального серого вещества у обезьян вызывает эмоциональные проявления, которые показывают, что такие раздражения воспринимаются как «наказание». О том, что перед нами истинное переживание неприятного аффекта, а не просто «псевдоаффективное» поведение, свидетельствует тот факт, что после нескольких опытов с раздражением этих областей животные обнаруживают признаки беспокойства, попадая в помещение, .где ставится опыт. Несомненно, что у них возникает «отрицательный мотив» вследствие сохранения в памяти неприятного опыта. Интересно, что вызываемые таким раздражением у кошек и обезьян реакции, 'подобные проявлениям ужаса, сохраняются без признаков утомления в течение всего периода раздражения (15 мин). То же относится и к раздражению покрышки, вызывающему несколько иной вид поведения. «Ручная и дружелюбная кошка рычала и набрасывалась на любой приближающийся предмет в течение 15 мин, пока производилось раздражение покрышки; как только раздражение прекращалось, восстанавливалось исходное состояние покоя» [177]. Такое поведение
ПСИХИЧЕСКИЕ ПРОЦЕССЫ С ФИЗИОЛОГИЧЕСКОЙ ТОЧКИ ЗРЕНИЯ 239
представляет резкий контраст двигательным реакциям, возникающим при раздражении коры, когда в течение нескольких секунд наступает выраженное утомление. Подобно медленно адаптирующимся болевым окончаниям периферических чувствительных нервов, эти центральные механизмы, которые также помогают избежать возможного повреждения, относятся к числу тех, активность которых сохраняется, пока не исчезает «опасность».
В литературе описаны реакции на раздражение подкорковых структур, которые явно указывают на участие последних в отрицательной мотивации. Реакции ярости, ужаса, нападения, защиты и т. п. почти несомненно со-1тровождаются изменениями мотивации, хотя в основном об этом приходится заключать по наблюдаемому поведению. Выраженные аффективные реакции непарко-тизированных кошек и обезьян удалось вызвать электрическим раздражением лимбической системы [23]. Организованные аффективные реакции были вызваны как электрическим, так и химическим (холинэргическим) раздражением лимбической части лобно-височной области у бодрствующих кошек и обезьян [93]. Эти реакции заключались в нападении и защите с соответствующими голосовыми реакциями.
Раздражение миндалевидного ядра у кошек вызывало интенсивные реакции ярости [176]. Из этих результатов можно сделать вывод, что данное раздражение возбуждает систему, активность которой чрезвычайно неприятна для животных, т. е создает отрицательную мотивацию.
Сходным образом существование областей отрицательной мотивации, или «наказания», было доказано у обезьян [576]. Раздражение этих областей вызывало реакции, указывающие на эмоции, варьирующие от незначительного до панического страха в зависимости от интенсивности раздражения. Следует отметить, что, если условия опыта позволяют обезьяне выключать раздражитель, она обычно делает это до того, как появятся внешние признаки страха. Если применяется сильный раздражитель, то животное делает отчаянные попытки избегнуть его. Вряд ли можно сомневаться в том, что
ГЛАВА IX
если бы обезьяна могла говорить, она бы выразила словами свое резко отрицательное отношение к условиям опыта и желание избегнуть его, короче говоря, засвидетельствовала бы наличие отрицательной мотивации. Суммируя, следует отметить наличие достаточно убедительных данных о том, что возбуждение одних областей мозга может служить как бы «поощрением» для животного, а возбуждение других — «наказанием». Правда, в этом вопросе следует проявить некоторую осторожность в свете одной из последних работ [115], в которой было показано, что кошку можно обучить либо искать, либо избегать возможности раздражения одной и той же точки латерального гипоталамуса в зависимости от условий опыта. Этот факт производит особенно большое впечатление потому, что все животные, использованные в этих экспериментах, были отобраны на основании проявления у них типичной реакции «гипотала-мической ярости» (а также беспорядочных движений) при таком раздражении. По нашему мнению, эти результаты не обесценивают представлений о существовании первичных систем поощрения и наказания в мозге, хотя Браун и Коэн [115] считают натяжкой всякие предположения о том, что самому по себе раздражению латерального гипоталамуса присущи отрицательные или «наказывающие» свойства, и рассматривают получаемый эффект как «ненаправленный» фактор мотивации. Возможно, однако, и другое объяснение, а именно что первичное влияние такого раздражения действует как «наказание», поскольку при этом возникает безусловная реакция ярости, но обучение, т. е. возникновение связей с соответствующими условиями и обстановкой, может приводить к возникновению иного общего типа активности центральной нервной системы при раздражении первичной системы «наказания», так что она вторично приобретает свойства системы «поощрения».
Физиологические механизмы мотивации
Обзор работ, анализирующих физиологические механизмы мотивации, позволяет предположить, что мотивация в основном определяется сравнительной актив-
ПСИХИЧЕСКИЕ ПРОЦЕССЫ С ФИЗИОЛОГИЧЕСКОЙ ТОЧКИ ЗРЕНИЯ 241
ностыо систем «поощрения» и «наказания», как описано в предыдущем разделе. Активность этих систем, очевидно, зависит от общей реакции активации, осуществляемой главным образом через ретикуло-гипоталами-ческую активирующую систему. Степень активности этой" системы частично определяется гуморальными воздействиями, а частично — нервными импульсами, которые она получает от висцеральной и соматической афферентных систем, а также от корковых и подкорковых структур, лежащих рострально от нее. «Поощряющий» .или «наказывающий» характер конечного эффекта активации, очевидно, определяется алгебраической суммой нервных и гуморальных влияний, опосредуемых более специализированными системами, такими, как система «голода — насыщения». В частности, для этой «специфической системы» можно указать определенные области гипоталамуса, имеющие особое значение для голода и насыщения. Остается неясным основной вопрос: каков механизм удовлетворения при других, биогенных влечениях? Следует доказать, достигается ли это во всех случаях активацией реципрокной «системы насыщения (поощрения)» или в некоторых случаях это связано1 со «снижением влечения». Фактически до сих пор не ясно, в какой мере «Центр насыщения», играющий важную роль в питании, может создавать рецип-рокное торможение «центра голода». Однако совершенно ясно, что потребность, как таковая, совсем не то же самое, что влечение. Потребность может существовать и быть даже очень высокой, но влечение не возникает без общей активации центральной нервной системы. Л в отсутствие активности соответствующей специфической системы поведение животного не будет направлено в нужную сторону. Так, если разрушить «центр голода» в гипоталамусе, крыса может погибнуть голодной смертью, не ощущая голода и не пытаясь найти пищу. Ясно также, что простое осуществление биологической потребности не обязательно устраняет влечение, ведущее к ее удовлетворению. Крыса с разрушенными центрами насыщения продолжает есть длительное время после того, как потребность в пище удовлетворена. Таким образом, потребность и влечение — это разные вещи. Вызовет ли
242 глава ix
потребность соответствующее влечение, зависит от того, каким образом потребность влияет на деятельность мозга.
Если учесть колоссальный диапазон эмоциональных оттенков, которые могут служить мотивами поведения, и огромное многообразие раздражителей, влияющих на эмоциональное состояние, то .постулат, согласно которому мотивация определяется равновесием между активностью систем «наказания» и «поощрения» в мозге, может показаться наивным. В связи с этим Мак-Лин [604] утверждает, что все известные эмоции следует рассматривать с точки зрения сохранения особи и сохранения вида и что они оказываются отрицательными или положительными в зависимости от того, связаны ли они с угрозой существованию или с устранением такой угрозы. Аналогичным образом Лилли [576] выдвигает весьма дискуссионное утверждение, что возникновение и повторение всех видов (двигательной) активности само по себе или обладает свойством внутреннего поощрения («старт-эффект») и (или) приводит к прекращению внутреннего «наказания» («егоп-эффект»). Он подчеркивает, что под словом «внутренний» он подразумевает «впутримозговой» и имеет в виду основные виды специфической нейрофизиологической активности, когда говорит о «поощрении и наказании».
Исследования, направленные на изучение мотивации — проблемы первостепенной важности для психологии, психиатрии, социологии, а фактически для самого существования человека на нашей планете,— очевидно, окажутся весьма плодотворными, если они будут исходить из этой гипотезы. Такие исследования уже были начаты, и полученные к настоящему времени результаты следует признать многообещающими.
Общие и специфические влечения
Мы указали ранее, что рассматриваем влечение как состоящее из внутреннего состояния и процессов, вызывающих и направляющих акты поведения. Эти состояния и процессы могут рассматриваться в свете общих
ПСИХИЧЕСКИЕ ПРОЦЕССЫ С ФИЗИОЛОГИЧЕСКОЙ ТОЧКИ ЗРЕНИЯ 243
и специфических влечений. Можно сказать, что состояние общего влечения возникает, когда уровень возбуждения центральной нервной системы достаточно высок для обеспечения готовности организма к деятельности. Это необходимо для любого мотивированного поведения. Специфические влечения изменяют активность определенных частей центральной нервной системы. Совпадение специфического и общего влечения создает активацию, которая вызывает и направляет поведение организма в целом.
Механизмы общего влечения
Кр-ид, Шеррингтон и др. [159] писали о «центральном состоянии возбуждения» и анализировали основные механизмы, посредством которых оно усиливает и ослабляет рефлекторные акты. Хотя природа центрального состояния возбуждения была неясной, предполагалось, что скорее всего это «специфическое проявление местного состояния возбуждения», которое, как было показано, связано с частичной деполяризацией периферических нервных волокон.
Морган [662] сформулировал физиологическую теорию влечения, в которой подчеркнул «центральное состояние мотивации», которое он рассматривает как синоним влечения, но пользуется этим термином, чтобы подчеркнуть локализацию истинных влечений, или состояний мотивации, в центральной нервной системе и чтобы снизить значение «периферических влечений», которые постулировались в то время в большом числе. Его «центральное состояние мотивации» немногим отличается от шеррингтоновского «центрального состояния возбуждения». Он признает, что мотивация меняется, если активность той или другой системы центров и путей повышается и становится выше остальных. Он считал, что диапазон мотивированного поведения определяется, уровнем центрального состояния мотивации.
Анализируя факторы, участвующие в половом повелении, Бич [47] также указывает на существование центрального механизма активации, порог которого понижается половыми гормонами. Такое гормональное
ГЛАВА IX
влияние «возможно, ограничивается подкорковыми центрами и низшими отделами головного и спинного мозга». Однако он указывает, что корковая часть механизма активации дополняет или усиливает влияние половых гормонов на низшие центры и в некоторых случаях может играть настолько важную роль в механизме активации, что половое поведение может осуществляться и в отсутствие половых гормонов. Бели и кортикальный, и гормональный компоненты отсутствуют, «механизм активации» полового поведения не действует и оно не осуществляется.
Гипоталамус и мотивация
В опубликованном в 1954 г. обзоре литературы, касающейся физиологических механизмов мотивации, Стеллар [864] сформулировал теорию, которую можно назвать «гилоталамической теорией мотивации». Ее основное положение заключается в том, что интенсивность реакций, связанных с мотивированным поведением, прямо пропорциональна активности определенных возбужденных центров' гипоталамуса. Таким образом, эта теория утверждает, что гипоталамус является местонахождением «центрального состояния мотивации», о котором писал Морган, и представляет собой тот «центральный нервный механизм», который Лешли объявил ответственным з<з организацию влечений.
Делл [181] дал обзор основных механизмов, посредством которых органические потребности трансформируются в акты поведения, и сделал некоторые убедительные замечания относительно общего влечения. Например, он утверждает, что во всех последних исследованиях инстинктивного поведения, обзор которых он приводит, отмечается наличие неспецифического компонента, общего для всех форм такого поведения. Этим
* * * *
' Хотя Стеллар пользуется для удобства термином «центр», он признает, что ни один нервный механизм не функционирует Изолированно, и достаточно осторожен, чтобы подчеркнуть, что он не намеревается допустить жесткую локализацию функций в изолированных анатомических структурах.
ПСИХИЧЕСКИЕ ПРОЦЕССЫ С ФИЗИОЛОГИЧЕСКОЙ ТОЧКИ ЗРЕНИЯ 245
компонентом служит повышенный уровень сенсорного и моторного возбуждения, которое «становится мотивом в буквальном смысле слова (производимого от слова tno-vere — двигаться, от которого происходит также и слово эмоция). Он подчеркивает, что при инстинктивном поведении этот неспецифический компонент может возникнуть задолго до специфических, что он представляет собой условие sine qua поп для проявления специфического поведения». При анализе случаев «кислородного голодания» Делл ссылается на экспериментальные данные, свидетельствующие о том, что низкое парциальное давление кислорода (р02) и высокое парциальное давление углекислоты (рС02) являются сильным раздражителем ретикулярной формации мозга. Гипоксия, по мнению Делла, возбуждает эту систему посредством двух механизмов: 1) путем усиления разрядов каротид-ных хеморецепторов и 2) путем прямого гуморального влияния продуктов обмена, возникающих при гипоксии. Гиперкапния является сильным раздражителем для передних областей ретикулярной формации, чувствительность которой к СОг оказалась столь же высокой, как и чувствительность классического дыхательного центра. Кроме перечисленных механизмов, «асфиксия вызывает мощный симпатический разряд, который, как известно, является сильным раздражителем восходящей и нисходящей облегчающей ретикулярной систем». «Симпатии», локально вырабатываемый в ретикулярной формации, также способствует тонической активности ее мезенце-фалической части [908]. При поиске пищи и пищевом поведении выделение адреналина под влиянием гипогликемии служит сильным раздражителем активирующей ретикулярной системы [181]. Было высказано предположение, что при всех этих формах «инстинктивного» поведения существует последовательность явлений, которая начинается с обменных сдвигов и продолжается процессами, при которых прежде всего используются запасы питательных веществ в организме (таких, как гликоген). По мере постепенного истощения запасов состав внутренней среды все более и более отклоняется от нормы. Такое отклонение воздействует на активность ретикулярной системы или прямым гуморальным путем,
Дата добавления: 2014-12-10; просмотров: 1143;