СИНТЕЗ ВЫСШИХ ЖИРНЫХ КИСЛОТ
Субстратом синтеза ВЖК является ацетил-КоА.Однако,в ходе синтеза жирных кислот (ЖК) в каждом цикле удлинения используется не сам ацетил-КоА, а его производное - малонил-КоА.
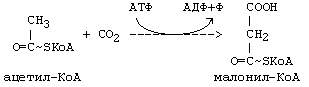
Эту реакцию катализирует фермент ацетил-КоА-карбоксилаза -ключевой фермент в мультиферментной системе синтеза ЖК. Активность фермента регулируется по типу отрицательной обратной связи. Ингибитором является продукт синтеза: ацил-КоА с длинной цепью (n=16) - пальмитоил-КоА. Активатором является цитрат. В состав небелковой части этого фермента входит витамин H (биотин).
В дальнейшем в ходе синтеза жирных кислот происходит поэтапное удлинение молекулы ацил-КоА на 2 углеродных атома за каждый этап за счет малонил-КоА, который в этом процессе удлинения теряет СО2.
После образования малонил-КоА основные реакции синтеза жирных кислот катализируются одним ферментом - синтетазой жирных кислот (фиксирован на мембранах эндоплазматического ретикулума). Синтетаза жирных кислот содержит 7 активных центров и АПБ (ацилпереносящий белок). Участок, связывающий малонил-КоА, содержит небелковый компонент – витамин B3 (пантотеновую кислоту). Последовательность одного цикла реакций синтеза ВЖК прпедставлен на рис.45.
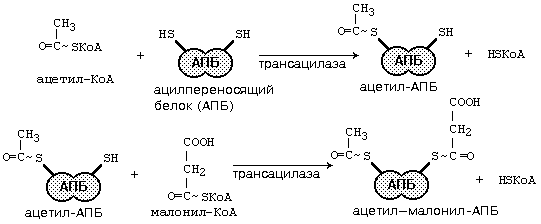
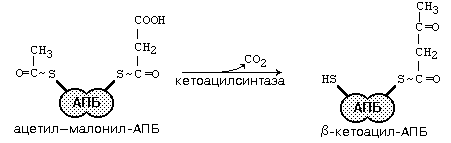
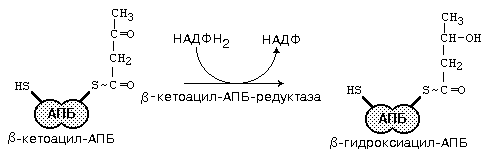
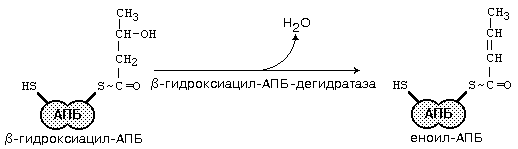
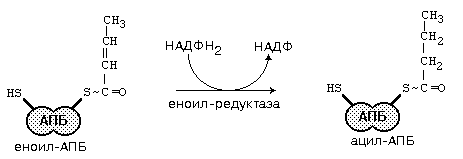
Рис.45. Реакции синтеза высших жирных кислот
После окончания цикла ацил-АПБ вступает в следующий цикл синтеза. К свободной SH-группе ацилпереносящего белка присоединяется новая молекула малонил-КоА. Затем происходит отщепление ацильного остатка, он переносится на малонильный остаток (с одновременным декарбоксилированием) и цикл реакций повторяется.
Таким образом, углеводородная цепочка будущей жирной кислоты постепенно растет (за каждый цикл – на два углеродных атома). Это происходит до момента, пока она не удлинится до 16 углеродных атомов (в случае синтеза пальмитиновой кислоты) или более (синтез других жирных кислот). Вслед за этим происходит тиолиз и образуется в готовом виде активная форма жирной кислоты – ацил-КоА.
Для нормального течения синтеза высших жирных кислот необходимы следующие условия:
1) Поступление углеводов, при окислении которых образуются необходимые субстраты и НАДФН2.
2) Высокий энергетический заряд клетки – высокое содержание АТФ, которое обеспечивает выход цитрата из митохондрий в цитоплазму.
Сравнительная характеристика b-окисления и синтеза высших жирных кислот:
1. b-окисление протекает в митохондриях, а синтез жирных кислот протекает в цитоплазме на мембранах эндоплазматического ретикулума. Однако, образовавшийся в митохондриях ацетил-КоА через мембраны сам проходить не может. Поэтому существуют механизмы транспорта ацетил-КоА из митохондрий в цитоплазму с участием ферментов цикла Кребса (рис.46).

Рис.46. Механизм транспорта ацетил-КоА из митохондрий в цитоплазму.
Ключевыми ферментами ЦТК являются цитратсинтаза и изоцитратдегидрогеназа. Основные аллостерические регуляторы этих ферментов - это АТФ и АДФ. Если в клетке много АТФ, то АТФ выступает как ингибитор этих ключевых ферментов. Однако изоцитратдегидрогеназа угнетается АТФ сильнее, чем цитратсинтетаза. Это приводит к накоплению цитрата и изоцитрата в матриксе митохондрии. При накоплении цитрат выходит из митохондрии в цитоплазму. В цитоплазме есть фермент цитратлиаза. Этот фермент расщепляет цитрат на ЩУК и ацетил-КоА.
Таким образом, условием для выхода ацетил-КоА из митохондрии в цитоплазму является хорошее обеспечение клетки АТФ. Если АТФ в клетке мало, то ацетил-КоА расщепляется до СО2 и Н2О.
2. В ходе b-окисления промежуточные продукты связаны с HS-КоА, а при синтезе жирных кислот промежуточные продукты связаны с особым ацил-переносящим белком (АПБ). Это сложный белок. Его небелковая часть похожа по строению на КоА и состоит из тиоэтиламина, пантотеновой кислоты (витамин В3) и фосфата.
3. При b-окислении в качестве окислителя используются НАД и ФАД. При синтезе ЖК нужен восстановитель - используется НАДФ*Н2.
В клетке существует 2 основных источника НАДФ*Н2 для синтеза жирных кислот:
а) пентозофосфатный путь распада углеводов;
б) в реакции окисления малата:
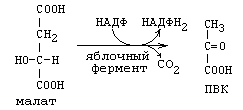
Эта реакция протекает в цитоплазме и катализируется ферментом малатдегидрогеназой.
Таким образом, обмен углеводов и обмен жиров очень тесно связаны. Углеводы легко могут превращаться в жиры, а вот превращение жиров в углеводы невозможно, так как ацетил-КоА не может превращаться в пируват.
Дата добавления: 2017-09-19; просмотров: 6138;