УТИЛИЗАЦИЯ КЕТОНОВЫХ ТЕЛ
Происходит в митохондриях (кроме клеток печени).
β-гидроксибутират превращается в ацетоацетат, а ацетоацетат вступает в реакцию с промежуточным продуктом ЦТК - сукцинил-КоА.
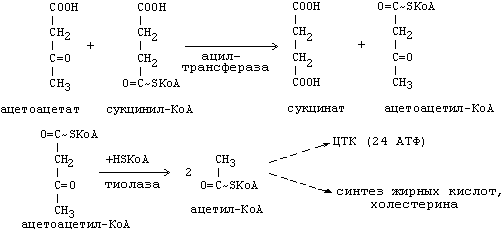
Пути использования образовавшегося из кетоновых тел ацетил-КоА зависят от функционального состояния клетки и ее специфики.
Однако чаще всего ацетил-КоА используется в ЦТК для получения энергии.
В норме процессы синтеза и использования кетоновых тел уравновешены, поэтому концентрация кетоновых тел в крови и в тканях обычно очень низка, и составляет 0.12 - 0.30 ммоль/л.
Однако при общем или при углеводном голодании может нарушаться баланс между образованием и утилизацией кетоновых тел. Это связано с тем, что скорость образования кетоновых тел зависит от скорости b-окисления жирных кислот в печени, а процесс b-окисления ускоряется при усилении липолиза (распада жира) в жировой ткани. Усиление липолиза может происходить под действием гормона адреналина, при мышечной работе, при голодании, при недостатке инсулина (сахарный диабет). В дальнейшем при усилении липолиза увеличивается скорость утилизации кетоновых тел, которые являются важными источниками энергии.
МЕТАБОЛИЗМ ХОЛЕСТЕРИНА В ОРГАНИЗМЕ. НАРУШЕНИЕ ОБМЕНА ЛИПИДОВ: АТЕРОСКЛЕРОЗ, ОЖИРЕНИЕ.
Холестерол – стероид, характерный только для животных организмов. Основное место его образования в организме человека – печень, где синтезируется 50% холестерола, в тонком кишечнике его образуется 15-20%, остальное количество синтезируется в коже, коре надпочечников и половых железах (рис.48).
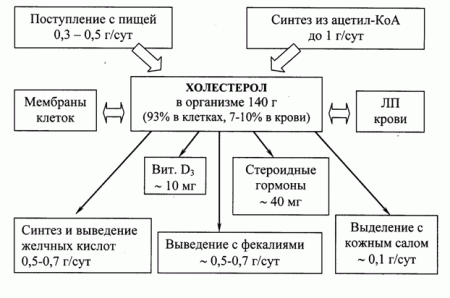
Рис.48. Пути поступления и выведения холестерола в организме.
Пищевой холестерин транспортируется хиломикронами и попадает в печень. Поэтому печень является для тканей источником и пищевого холестерина (попавшего туда в составе хиломикронов), и эндогенного холестерина.
Синтез холестерина протекает в основном в печени на мембранах эндоплазматического ретикулума гепатоцитов. Этот холестерин - эндогенный. Происходит постоянный транспорт холестерина из печени в ткани. Для построения мембран используется также пищевой (экзогенный) холестерин. Ключевой фермент биосинтеза холестерина - ГМГ-редуктаза (β-гидрокси-β-метил-глутарил-КоА-редуктаза). Этот фермент ингибируется по принципу отрицательной обратной связи конечным продуктом – холестерином (рис.49).
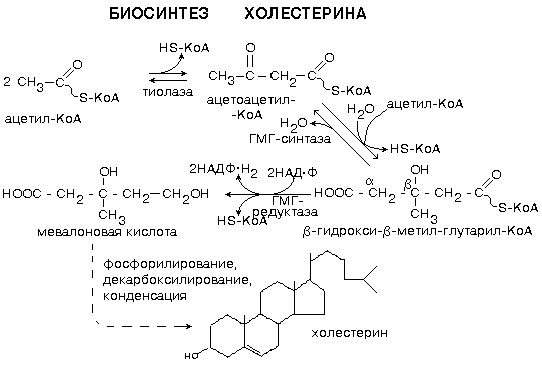
Рис.49. Схема синтеза холестерина
В печени синтезируются и затем попадают в кровь ЛОНП - липопротеины очень низкой плотности (состоят на 75% из холестерина), а также ЛПНП - липопротеины низкой плотности, в состав которых входит апобелок апоВ100.
Почти во всех клетках имеются рецепторы для апоВ100. Поэтому ЛПНП фиксируются на поверхности клеток. При этом наблюдается переход холестерина в клеточные мембраны. Поэтому ЛПНП способны снабжать холестерином клетки тканей.
Помимо этого, происходит освобождение холестерина из тканей и транспорт его в печень. Транспортируют холестерин из тканей в печень липопротеины высокой плотности (ЛПВП). Они содержат очень мало липидов и много белка. Синтез ЛПВП протекает в печени. Частицы ЛПВП имеют форму диска, и в их составе находятся апобелки апоА, апоС и апоЕ. В кровеносном русле к ЛПНП присоединяется белок-фермент лецитинхолестеринацилтрансфераза (ЛХАТ).
АпоС и апоЕ могут переходить от ЛПВП на хиломикроны или ЛОНП. Поэтому ЛПВП являются донорами апоЕ и апоС. АпоА является активатором ЛХАТ.
ЛХАТ катализирует следующую реакцию:
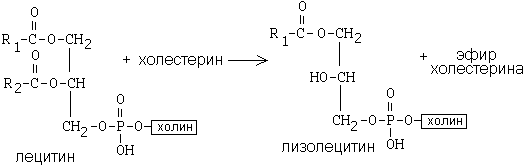
Это реакция переноса жирной кислоты из положения R2 на холестерин.
Реакция является очень важной, потому что образующийся эфир холестерина является очень гидрофобным веществом и сразу переходит в ядро ЛПВП - так при контакте с мембранами клеток ЛПВП удаляют из них избыток холестерина. Дальше ЛПВП идут в печень, там разрушаются, и избыток холестерина удаляется из организма.
К патологиям обмена липидов относятся: ожирение, атеросклероз, гиперлипопротеинемия, болезнь Тея-Сакса – при которой наблюдается накопление значительного количества липидов в лизосомах.
Дата добавления: 2017-09-19; просмотров: 1702;