Тема 9: Генетика пола
Пол – это совокупность признаков и свойств организма, обеспечивающих его участие в воспроизводстве потомства и передаче наследственной информации за счет образования гамет.
Обычно признаки, по которым отличаются особи разных полов, делят на первичные и вторичные половые признаки. К первичным относят те морфологические и физиологические особенности организма, которые обеспечивают образование гамет и соединение их в процессе оплодотворения. К числу их относятся, например, гонады, половые пути и наружные гениталии у высших животных, андроцей и гинецей у высших растений. К вторичным половым признакам относят признаки и свойства организма, непосредственно не обеспечивающие процессы гаметогенеза, спаривания и оплодотворения, но играющие некоторую вспомогательную роль в половом размножении. К ним относятся особенности строения плавников у рыб, оперения у птиц, грудных желез у млекопитающих, длины междоузлий и сроков цветения у высших растений и др.
Из числа вторично-половых признаков выделяют иногда ограниченные полом признаки. Генетическую информацию о них несут все особи, но проявляются они только у одного пола. Так, быки несут гены, определяющие молочность, петухи - гены, определяющие яйценоскость, но действие их у самцов не проявляется.
Следует отличать также признаки, зависимые от пола. К числу зависимых от пола признаков относятся такие признаки, характер доминирования которых в гетерозиготе зависит от пола особи. Так, доминантные гомозиготные овцы (НН) рогаты, а гомозиготные рецессивы (hh) безроги, независимо от пола. Однако гетерозиготные (Hh) самцы рогатые, а самки безрогие. Аналогично наследуется раннее облысение у человека. Доминирование в таких случаях определяется количеством мужских и женских половых гормонов в крови. Знание закономерности наследования признаков, зависимых от пола, позволяет еще глубже понять сущность явления доминирования и подойти к практическому управлению доминированием.
Различия особей мужского и женского пола - половой диморфизм встречается у некоторых низших и многих высших растений и животных. Среди высших растений около 5000 видов двудомны. У высших животных обоеполые (гермафродитные) формы в норме не встречаются. Во всех случаях отличия касаются морфологических, биохимических, физиологических признаков, экологии, поведения у животных и т. д. Однако на низших ступенях эволюции у одноклеточных организмов отличия особей, вступающих в копуляцию, могут быть невидимы и касаться лишь физиологических особенностей, поэтому и пол их обозначают условно как « + пол» или «-пол». Различия между этими двумя типами часто носят количественный характер, так что у многих одноклеточных водорослей (хламидомонады, например), некоторых низших грибов или инфузорий удается выделить несколько линий, внутри которых копуляция клеток невозможна, а между линиями она осуществляется. Удалось сгруппировать многочисленные линии в « + » и «-» типы, но оказалось, что и внутри типа копуляция клеток разных линий иногда происходит. Следовательно, здесь можно говорить об относительной сексуальности.
Относительная сексуальность особенно хорошо изучена у хламидомонад М. Гартманом и Ф. Мевусом. Они показали, что гаметы даже одного пола, но с разной половой валентностью способны воспроизводить потомство половым путем. Половые различия гамет связаны с гормоноподобными веществами (гамонами).
Научное обоснование полового диморфизма стало возможным благодаря развитию цитологии и генетики. У всех раздельнополых организмов были обнаружены хромосомы, одинаковые у обоих полов, и хромосомы, по которым, мужской и женский пол отличаются друг от друга. Хромосомы одинаковые для женских и мужских особей назвали аутосомами,а отличающиеся- половыми, или гетерохромосомами (гоносомами). Например, в клетке человека содержится 46 хромосом или 23 пары: 22 пары аутосом и 1 пара половых хромосом.
Соотношение полов у раздельнополых организмов близко 1 : 1, т.е. самцы и самки встречаются одинаково часто. Еще Мендель обратил внимание, что такое расщепление характерно для скрещивания особей: Аа х аа. Из это следует, что один из полов должен быть гомозиготным, а другой – гетерозиготным. Пол, образующий одинаковые в отношении определения пола гаметы, называют гомогаметным, а пол, образующий разные гаметы – гетерогаметным. В случае гетерогаметности мужских особей половые хромосомы обозначают как X- и Y-хромосомы. В случае гетерогаметности женских особей самка обозначается ZW, самец – ZZ.
Изучение половых хромосом показало, что они отличаются от аутосом и генетически и цитологически. В них содержится меньшее число аллелей, они богаты гетерохроматином, редупликация их происходит асинхронно с аутосомами, а у гомогаметного пола одна из Х хромосом репродуцируется позже остальных. В мейозе гоносомы часто спирализованы (гетеропиктоз). Половые хромосомы у гетерогаметного пола не конъюгируют или конъюгируют лишь частично, что указывает на частичную гомологичность (гемизиготность). В этом случае проявляется рецессивный ген, имеющийся в генотипе в единственном числе. Так X-хромосома может содержать ген, вызывающий гемофилию (пониженную свертываемость крови). Тогда все мужские особи, получившие эту хромосому, будут страдать этим заболеванием, так как Y-хромосома не содержит доминантного аллеля.
Анализ половых хромосом у различных организмов позволил сформулировать хромосомную теорию определения пола и показал, что существует 5 типов хромосомного определения пола:
1) ♀ XX, ♂ ХУ - характерен для млекопитающих (в том числе и для человека), червей, ракообразных, большинства насекомых (в том числе для дрозофил), большинства земноводных, некоторых рыб (рис. 4).
2) ♀ ZW, ♂ ZZ - характерен для птиц, пресмыкающихся, некоторых земноводных и рыб, некоторых насекомых (чешуекрылые).
3) ♀ XX, ♂ Х0 - (0 обозначает отсутствие хромосом) встречается у некоторых насекомых (прямокрылые).
4) ♀ Z0, ♂ ZZ - встречается у некоторых насекомых (равнокрылые).
5) гапло-диплоидный тип (♀ 2n, ♂ n) встречается, например, у пчел и муравьев: самцы развиваются из неоплодотворенных гаплоидных яйцеклеток (партеногенез), самки - из оплодотворенных диплоидных.
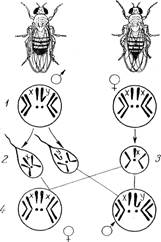
Рисунок 4. Схема определения пола у дрозофилы. Хромосомные наборы: 1 - соматических клеток; 2-— сперматозоидов; 3 - яйцеклеток; 4 - потомства.
Таким образом, хромосомный механизм определения пола является идеальным саморегулирующимся механизмом. В результате оплодотворения возникает равное количество самцов и самок.
Однако, иногда встречаются такие явления, которые как будто специально созданы природой для проверки правильности теории. В отношении хромосомной теории примером может служить явление гинандроморфизма. Организмы, совмещающие в себе части тела разных полов - мужского и женского, называют гинандроморфами (гин -♀ , андр - ♂ ). Гинандроморфы существуют у тех видов, у которых четко выражен половой диморфизм (насекомые, птицы, человек), но встречаются они редко.
При латеральном гинандроморфизме, например у дрозофилы, одна половина тела имеет признаки женского пола, а другая - мужского (рис. 5). Как может возникнуть такой организм? Цитологические исследования показывают, что ткани гинандроморфа химерны: женская половина несет две Х-хромосомы, а мужская - одну.
На приведенном рисунке показан случай, когда у гинандроморфа рецессивный, сцепленный с полом ген white проявился на мужской стороне тела и не проявился на женской. Почему это так?
У гинандроморфа, возникшего из зиготы w+w, при первом делении дробления в силу каких-то необычных условий одна из Х-хромосом, несущая ген w+, в одной из дочерних клеток (бластомеров) утрачивается. Тогда две дочерние клетки окажутся неодинаковыми в отношении Х-хромосом:
одна w , а вторая w.
w+
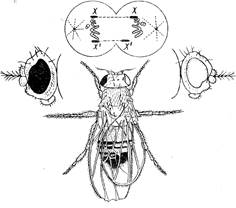
Рисунок 5. Гинандроморф дрозофилы. Вверху - предполагаемая схема возникновения; X - элиминируется.
Половина тела мухи, развившаяся из первой клетки, окажется женской и с красным глазом, а из второй разовьется половина тела с признаками мужского пола и с белым глазом, поскольку рецессивный ген w, содержащийся в единственной Х-хромосоме, будет в гемизиготном состоянии.
Таким образом, и цитологический, и генетический анализ показывает, что в данном случае причиной гинандроморфизма может быть элиминация одной из Х-хромосом.
Кроме этого типа гинандроморфизма, который можно назвать монозиготным, известен также дизиготический гинандроморфизм. Он обнаружен у бабочек - Abraxas, тутового шелкопряда и у дрозофилы. Например, иногда в яйцеклетке тутового шелкопряда (самка гетерогаметна) образуются два женских пронуклеуса, один из которых кроме аутосом (обозначим их А) содержит Х-хромосому (Х + А), а другой — У + А. При полиспермии оба пронуклеуса будут оплодотворены разными спермиями, тогда в одном из бластомеров будет ХХ + АА, а в другом — ХУ+АА. Это и приведет к развитию дизиготного гинандроморфа. Аналогично может возникать гинандроморф у дрозофилы, только здесь различия между бластомерами получаются за счет разных сперматозоидов (самцы гетерогаметны).
Исключения из хромосомной теории определения пола. По мере накопления фактов хромосомная теория определения пола не только находила подтверждение, но и встречала некоторые трудности. Оставался открытым вопрос о том, не являются ли половые хромосомы индикаторами пола, вторичнополовыми признаками?
Анализ исключительных особей у дрозофил, которые были получены в опытах Бриджеса, как результат нерасхождения половых хромосом показал, что особи, имеющие, кроме аутосом, ХХУ-хромосомы (ХХУ + АА), являются самками, а особи ХО + АА—самцами. Эти факты убедительно говорили о том, что половые хромосомы отнюдь не индикаторы пола. Но как же они определяют пол, если особи ХУ+АА и ХО+АА являются самцами, а ХХ+АА и ХХУ+АА самками? Очевидно, дело обстоит не так просто, как это казалось вначале.
Дата добавления: 2016-04-22; просмотров: 1424;