Тема 8: Сцепленное наследование. Генетические карты.
При скрещивании особей, различающихся двумя парами признаков (АВ и ав), согласно закону независимого наследования признаков, гибрид АаВв будет производить четыре сорта гамет: АВ, Ав, аВ, ав. Причем, эти гаметы должны производиться в равных количествах. Теоретически при анализирующем скрещивании дигибрида АаВв с аавв,расщепление генотипов должно составлять 1:1:1:1.
Однако, уже в начале 20-го века, было накоплено множество фактов, что в некоторых случаях у потомков, доля родительских генотипов оказывается выше предполагаемой, т.е. процент рекомбинантных типов наблюдается ниже ожидаемого, а иногда новые комбинации признаков (Ав и аВ) вовсе отсутствуют.
Это явление было названо сцеплением генов. Изучением сцепления признаков занимались Стертенвант. И Т.Х. Морган с учениками в начале 20-х годов XX в. Объектом их исследований являлась плодовая мушка дрозофила (срок ее жизни невелик, и за год можно получить несколько десятков поколений, ее кариотип составляют всего четыре пары хромосом). Было выяснено, что закон независимого наследования генов справедлив только для генов, расположенных в разных хромосомах. Гены, лежащие в одной хромосоме наследуются преимущественно вместе и называются группой сцепления. Количество групп сцепления организмов соответствует количеству хромосом гаплоидного набора клетки. При локализации аллелей в половых хромосомах наблюдается наследование признаков, сцепленным с полом. В половых хромосомах могут находиться гены, не имеющие отношения к развитию половых признаков.
Однако, в некотором проценте случаев сцепление все же может нарушаться. Причина нарушения сцепления – кроссинговер, наблюдающийся в профазе I мейотического деления. Кроссинговер приводит к нарушению групп сцепления и генетической рекомбинации.
Гаметы с хромосомами, претерпевшими кроссинговер, называют кроссоверными, а не претерпевшие - некроссоверными. Соответственно организмы, возникшие от сочетания кроссоверных гамет гибрида с гаметами анализатора, называют кроссоверами или рекомбинантами, а возникшие от сочетания некроссоверных гамет с гаметами анализатора - некроссоверными или нерекомбинантными.
Явление кроссинговера, так же как и сцепление, оказалось общим для всех животных, растений и микроорганизмов. Наличие обмена идентичными участками между гомологичными хромосомами обеспечивает рекомбинацию генов и тем самым значительно увеличивает роль комбинативной изменчивости в эволюции.
Генетический анализ кроссинговера. Какими же генетическими методами можно отличить явление сцепленного наследования от независимого комбинирования генов?
О перекресте хромосом можно судить на основе учета частоты возникновения организмов с новым сочетанием признаков. Классическим примеров проявления кроссинговера являются опыты Моргана по скрещиванию мух с определенным фенотипом. Правила записи независимых и сцепленных генов следующие: гены находящиеся в разных парах хромосом, например, генотип дигетерозиготы записывается так: АВ
а b
Если же гены находятся в одной паре гомологичных хромосом, формула видоизменяется:
АВ
аb
При этом аллели одного гена (Аа и Вb), находящиеся в гомологичных хромосомах, пишутся строго одна под другой.
У дрозофилы рецессивный ген черной окраски тела обозначается в, а его доминантная аллель, определяющая дикую серую окраску, -b +, ген рудиментарных крыльев - vg, нормальных vg+. При скрещивании мух, различающихся по двум парам сцепленных признаков, - серых с рудиментарными крыльями b+ vg и черных с нормальными крыльями b vg+
b+ vg b vg+
Гибриды F1 b+ vg
b vg+ по фенотипу - серые с нормальными крыльями.
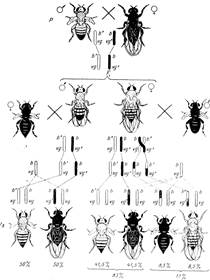
На рисунке 1 представлены два анализирующих скрещивания: в одном из которых дигетерозиготой является самец, в другом - самка. Если гибридные самцы скрещиваются с самками, гомозиготными по обоим рецессивным генам,
♀ b vg х ♂ b+ vg
b vg b vg+
то в потомстве получается расщепление в отношении 1 серотелая с рудиментарными крыльями : 1 чернотелая с нормальными крыльями, т.е. 50% : 50% .
1 2
Рисунок 1. Наследование сцепленных признаков у дрозофилы: 1- в случае отсутствия кроссинговера (гетерозиготный самец F1); 2 –в случае наличия кроссинговера (гетерозиготная самка F1); В Fb изображены только самки, т.к. самцы имеют те же фенотипы, что и у родителей; b + - серая, b - темная окраска тела, vg+ - нормальные крылья, vg – зачаточные крылья.
Такое расщепление показывает, что данная дигетерозигота образует только два сорта гамет b+vg и bvg+ вместо четырех, причем сочетание генов в гаметах самца соответствует тому, которое было у его родителей. Исходя из указанного расщепления, следует предположить, что у самца не происходит обмена участками гомологичных хромосом. В дальнейшем выяснилось, что у самцов дрозофилы действительно как в аутосомах, так и в половых хромосомах в норме не происходит кроссинговер. Поэтому при анализирующем скрещивании в потомстве появляются только две исходные родительские комбинации признаков в равных количествах, что указывает на полное сцепление генов, находящихся в одной паре гомологичных хромосом.
Может возникнуть предположение, что серая окраска тела и рудиментарные крылья, а также черное тело и нормальные крылья - это пары признаков, наследующихся вместе вследствие плейотропного действия одного гена. Однако если взять для анализа гетерозиготных самок, а не самцов, то в F1, наблюдается иное расщепление. Кроме родительских комбинаций признаков появляются новые типы - мухи с черным телом и рудиментарными крыльями, а также мухи с серым телом и нормальными крыльями. В этом скрещивании сцепление тех же генов нарушается за счет того, что гены в гомологичных хромосомах поменялись местами благодаря кроссинговеру.
Открытое Морганом сцепленное наследование признаков получило название закона сцепления Моргана: гены, локализованные в одной хромосоме, наследуются преимущественновместе.
При анализе расщепления в случае кроссинговера обращает на себя внимание определенное количественное отношение кроссоверных и некроссоверных классов. Обе исходные родительские комбинации признаков, образовавшиеся из некроссоверных гамет, оказываются в потомстве анализирующего скрещивания в равном количественном отношении. В указанном, опыте с дрозофилой тех и других особей было примерно по 41,5%. В сумме некроссоверные мухи составили 83% от общего числа потомков. Два кроссоверных класса по числу особей также одинаковы, и сумма их равна 17%.
Частота кроссинговера не зависит от аллельного состояния генов, участвующих в скрещивании. Если в качестве родителей использовать мух,
♀ b vg х ♂ b+ vg+
b vg b+vg +
то в анализирующем скрещивании кроссоверные b+vg и b vg+ и некроссоверные - b vg и b+vg+ особи появятся с той же частотой (соответственно 17 и 83%), что и в первом случае.
Результаты этих опытов показывают, что сцепление генов действительно существует, и лишь в известном проценте случаев оно нарушается вследствие кроссинговера. Отсюда и был сделан вывод, что между гомологичными хромосомами может осуществляться взаимный обмен идентичными участками, в результате чего гены, находящиеся в этих участках парных хромосом, перемещаются из одной гомологичной хромосомы в другую. Отсутствие перекреста (полное сцепление) между генами представляет исключение и известно лишь у гетерогаметного пола двух видов - дрозофилы и шелкопряда.
Наследование при плейотропном действии гена.Однако совместное наследование признаков при отсутствии рекомбинантов может быть обусловлено плейотропным действием одного гена. Например, у кроликов доминантный ген R определяет нормальную длину шерсти и прямые усы, а рецессивная аллель r - короткую шерсть и извитые усы. При анализирующем скрещивании гетерозиготы Rr любого пола с рецессивной гомозиготой rr в Fb будут появляться в отношении 1 : 1 нормальношерстные кролики с прямыми усами Rr и короткошерстные с извитыми усами rr . Таким образом, в данном случае наблюдается картина полного сцепления признаков за счет плейотропии, но не сцепления генов.
Морган выяснил, что в хромосоме может одновременно происходить несколько перекрестов. Кроссинговер, происходящий в одном месте называется – одинарным, в двух точках одновременно - двойным, в трех – тройным и т.д., т.е. он может быть множественным (рис. 2).
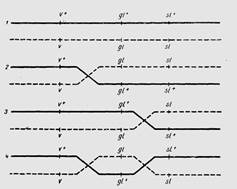
Рисунок 2. Рекомбинация сцепленных генов у кукурузы: 1 – некроссоверные хромосомы; 2- кроссинговер между генами v и gl; 3- кроссинговер между генами gl и sl; 4 – двойной кроссинговер между генами v-gl и gl-sl.
Установлено, что кроссинговер, происходящий в одном месте хромосомы, подавляет кроссинговер в близлежащих районах. Это явление получило название - интерференция.Величину интерференции измеряется отношением наблюдаемого числа двойных перекрестов к теоретически ожидаемому.
Частота кроссинговера имеет практическое значение для определения местоположения генов в хромосоме. Чем дальше друг от друга расположены гены, тем чаще между ними происходит кроссинговер, чем реже кроссинговер, тем ближе они находятся друг к другу. Величина кроссинговера измеряется в процентах и отражает силу сцепления генов в хромосоме: чем она больше, тем меньше сила сцепления.
На этом явлении основано построение генетических карт - определение последовательности расположения генов в хромосоме и примерного расстояния между ними.
В качестве условной единицы расстояния на карте для определения локализации генов в хромосоме, Стертевант предложил использовать частоту рекомбинации между ними:
Расстояние на карте = (Число рекомбинантов)/ (общее число потомков) х 100.
Одна единица карты (сантиморганида) соответствует 1 % рекомбинантных потомков.
Генетические карты составляют для каждой пары гомологичных хромосом. При составлении генетических карт указывается группа сцепления, полное или сокращенное название генов, расстояние в процентах от одного из концов хромосомы, принятого за нулевую точку; иногда обозначается место центромеры. Установлено, что длина генетической карты связана с величиной хромосомы.
Таким образом, генетическая карта представляет собой схему относительного расположения генов, находящихся в данной группе сцепления. Они составлены для некоторых наиболее изученных с генетической точки зрения объектов: дрозофилы, кукурузы, томатов, мыши, нейроспоры, кишечной палочки. В настоящее время составлены генетические карты и для человека.
Для того чтобы составить карты, необходимо изучить закономерности наследования большого числа генов. У дрозофилы, например, изучено более 500 генов, локализованных в 4-х группах сцепления, у кукурузы – более 400 генов. Локализованных в 10-ти группах сцепления и т. д. Карта хромосом кукурузы, приведенные в виде сокращенных схем (рис. 3), являются плодом огромного и систематического труда многих исследований.

Рисунок 3. Генетические карты хромосом кукурузы: I-Х - группы сцепления; цифры указывают расстояние между генами и одним из концов хромосомы (в единицах перекреста). Центромеры указаны кружком.
Названия генов: I: sr1 - полосатые листья; ga6 - гаметофитный фактор; ms17 - мужская стерильyjсть; ts2 - метелка с семенами; Р - окрашенный перикарп; zl - зиготическая леталь; as - асинапсис; hm - устойчивость к гельминтоспориозу; br1 - укороченные междоузлия; vg - зачаточные султаны; f1 - тонко исчерченные листья; аn1 - початок с тычинками; Кn - узловатые листья; gs1,- листья с зелеными полосами; Ts6 - метелка с семенами; bm2 - коричневатая средняя жилка листа.
Создание генетических карт позволяет предсказывать характер наследования признаков, гены которых нанесены на карту, а в селекционной работе облегчает подбор пар для скрещивания.
При изучении закономерностей рекомбинаций и составлении карт было установлено:
1. Индивидуальные расстояния между генами аддитивны, т.е. гены расположены последовательно.Если два гена А. В. Находятся на расстоянии 10 единиц друг от друга, а расстояние от В до С составляет 5 единиц, то расстояние между А и С составит 15 ед.
А…………………….В………..С
10 ед. 5 ед.
2. Расстояние между генами не зависит от типа аллелей, а зависит только от положения генных локусов,т.е. места в хромосоме, в котором находится ген данного признака. Все аллели данного гена всегда находятся в одном и том же локусе.
3. Не все пары генов, локализованных в одной хромосоме сцеплены, но максимальное число рекомбинантов не может превышать 50 %, т.к. такое соотношение гамет с родительскими и новыми сочетаниями генов обнаруживается при независимом наследовании.
Следовательно, расстояние между самыми крайними точками в пределах одной хромосомы не может составить более 50%. Однако, при рассмотрение генетических карт в группе сцепления кукурузы имеются гены, занимающего положения 62 и 107%. Это кажущееся несоответствие объясняется тем, что локализация генов осуществляется путем учета кроссинговера на коротких, последовательно взятых участках, а на карту наносится сумма величин кроссинговера, определенная для всех участков. Поэтому, общая длина генетической карты может значительно превышать полученную в эксперименте.
Дата добавления: 2016-04-22; просмотров: 3557;