Н-рефлекса при патологии
Н-рефлекс является аналогом Ахиллова рефлекса, однако афферентное раздражение при вызывании Н-рефлекса возникает не в мышечных рецепторах Ia волокон, как при ахилловом рефлексе, а на протяжении волокон в подколенной ямке (рис. 117).
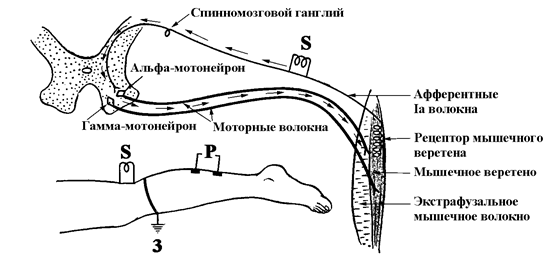
Рис. 117. Схема рефлекторной дуги Н-рефлекса (По Ст.Байкушеву и соавт., 1974. Адаптировано).
S – стимулирующие электроды,
Р – регистрирующие электроды,
З – заземляющий электрод.
Исследование Н-рефлекса позволяет судить о состоянии проводимости всей сегментарной дуги, включая сенсорные, двигательные волокна вне спинного мозга и интраспинальную часть, а также об уровне возбудимости мотонейронов.
В противоположность F‑волне, регистрируемой в большинстве мышц, Н-рефлекс вызывается в ограниченном числе мышц (m.soleus, m.gastrocnemius, flexor carpi radialis), имеющих отношение к антигравитационной нагрузке (мышцы голени) и являющихся преимущественно тоническими. Однако произвольная легкая активация мышц, приводя к повышению возбудимости периферических мотонейронов, способствует регистрации Н-рефлекса в большем числе мышц. Вместе с тем, этот прием активации мотонейронов приводит к незначительному повышению латентности Н-рефлекса, что необходимо учитывать при проведении сравнительных исследований.
Регистрация Н-рефлекса в большинстве мышц у новорожденных и детей до 1 года и последующее угасание его, с сохранением только в мышцах голени, свидетельствует о связи Н-рефлекса со зрелостью и сохранностью нисходящих тормозных влияний головного мозга на спинальный аппарат спинного мозга.
Максимальная активация Н-рефлекса происходит в условиях стимуляции нерва током средней интенсивности, позволяющим активировать низкопороговые Ia сенсорные волокна и оставлять при этой интенсивности стимуляции неактивированными моторные волокна нерва. Большая интенсивность стимуляции приводит к постепенному снижению и последующему исчезновению Н-рефлекса. Повышение интенсивности раздражения нерва активирует дополнительно моторные волокна нерва, что ведет к развитию тормозных процессов. Механизмы торможения Н-рефлекса при супрамаксимальной стимуляции могут реализовываться по нескольким путям:
· возникновение рефрактерной фазы возбудимости мотонейрона после его антидромной активации,
· коллизия – столкновение импульсов в моторных клетках, приходящих к ним по сенсорным волокнам ортодромно и по двигательным антидромно,
· активация клеток Реншоу при антидромном движении раздражающих импульсов по двигательным волокнам,
· активация одновременно с Ia волокнами Ib сенсорных волокон, идущих от сухожилий, которая приводит к постсинаптическому торможению альфа-мотонейронов (E.Pierrot-Deseilligny et al., 1981).
В отличии от F-волны Н-рефлекс более постоянен по латентности. В незначительной степени латентность Н-рефлекса возрастает при увеличении длительности раздражающего импульса до 0.5-1.0 мс (M.Panizza et al., 1989) и произвольной активации мышцы в период регистрации Н-рефлекса. Амплитуда Н-рефлекса достаточно изменчива и зависит от ряда условий.
1. Состояние произвольной активации мышцы – при напряжении мышцы амплитуда Н-рефлекса выше, в состоянии расслабления снижается.
2. Состояние пассивного растяжения мышцы – при растяжении мышцы (миотатическом рефлексе) амплитуда Н-рефлекса снижается.
3. Рефлекторная вибрационная активация мышцы (тоническое напряжение мышцы при приложении к сухожилию вибратора, генерирующего частоту 100-200 Гц амплитудой 0.5-1.0 мм), которая приводит к исчезновению Н-рефлекса у здоровых испытуемых (аналог миотатического рефлекса).
4. Частота стимуляции – при частой стимуляции нерва (более 1 раздражения в 10 секунд) амплитуда Н-рефлекса снижается, что связано с понижением возбудимости мотонейронов от предшествующего стимула.
На основании зависимости амплитуды Н-рефлекса от частоты стимуляции предложена методика оценки кривой восстановления возбудимости мотонейронов при парной стимуляции нерва с межстимульным интервалом в диапазоне от 5 до 1000 мс. В кривой восстановления возбудимости Н-рефлекса выделяют 3 периода торможения. Первичное торможение возникает в диапазоне 1-5 мс, вторичное торможение – в диапазоне 8-80 мс и период угнетения – в диапазоне 300-400 мс. Для последнего периода амплитуда второго (тестирующего) Н-рефлекса ниже первого (кондиционирующего) на 50% (рис. 118).
Основным механизмом торможения сегментарной возбудимости кондиционирующим импульсом является пресинаптическое торможение и тормозные влияния клеток Реншоу (В.С.Гурфинкель, 1955). Пресинаптическое торможение обусловлено тормозным влиянием интернейронов сегментарного аппарата спинного мозга на терминали сенсорных нейронов, моносинаптически связанные с моторными клетками передних рогов спинного мозга. Ориентировочную оценку пресинаптического торможения можно проводить усеченной методикой при парной стимуляции с межстимульным интервалом 330 мс (3 Гц).
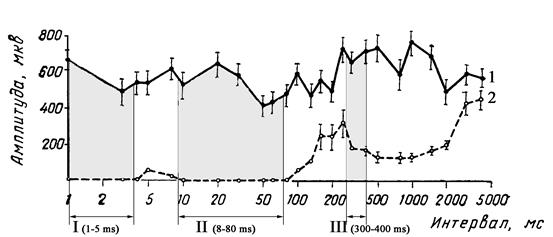
Рис. 118. Кривая восстановления Н-рефлекса в норме (По Ст.Байкушеву и соавт., 1974. Адаптировано).
1 – амплитуда Н-рефлекса при стимуляции околопороговым кондиционирующим стимулом,
2 – амплитуда Н-рефлекса при стимуляции тестирующим стимулом,
I – период первичного торможения,
II – период вторичного торможения,
III – период угнетения Н-рефлекса.
В норме раздражение афферентных волокон приводит к активации только части (наиболее возбудимой) мотонейронного пула в передних рогах спинного мозга с последующей потерей их рефлекторной активации при усилении интенсивности раздражения и развитии торможения. Эти особенности отличают Н-рефлекс от F‑волны, которая возникает только при максимальном раздражающем стимуле и имеет значительно меньшую амплитуду (табл. 29).
Таблица 29
Сравнение Н-рефлекса и F-волны
| Критерии отличия | Н-рефлекс | F-волна |
| Моносинаптический рефлекс | Возвратный разряд в моторных нейронах | |
| Интенсивность раздражающего стимула в сравнении с возбуждением М-ответа | Низкая (под- или надпороговая) | Высокая (супрамаксимальная) |
| Подавление ответа супрамаксимальным стимулом | Подавляется | Не подавляется |
| Постоянство латентности ответа | Относительно постоянная латентность | Колебание латентности от стимула к стимулу |
| Выраженность амплитуды по сравнению с М‑ответом | Большая (10-70%) | Низкая (1-5%) |
| Постоянство амплитуды ответа | Постоянна при редкой частоте стимуляции (реже чем 1 в 10 секунд) | Колебание амплитуды от стимула к стимулу |
| Форма ответа | Постоянна | Изменчива от стимула к стимулу |
Абсолютное значение амплитуды Н-рефлекса значительно варьирует, поэтому величина амплитуды нормируется по амплитуде моторного ответа. Процентное отношение максимальной амплитуда Н-рефлекса к максимальной амплитуде М-ответа является мерой сегментарной возбудимости мотонейронов (R.W.Angel, W.W.Hoffmann, 1963; Я.М.Коц, 1972). Определение сегментарной возбудимости мотонейронов спинного мозга для разных мышц голени – икроножной и камбаловидной - дает характеристику разным регуляторным механизмам - преимущественно церебральным или спинальным – соответственно. Это связано c функциональным различием этих мышц. Икроножная мышца в большей степени контролируется корой и содержит преимущественно быстрые двигательные единицы, а камбаловидная – медленные (Я.М.Коц, 1975).
Сопоставление Н-рефлекса с сухожильным (ахилловым) рефлексом (или Т-рефлексом) дает возможность оценить состояние гамма-моторной системы. Н‑рефлекс реализуется вне гамма-системы, сухожильный ахиллов рефлекс связан с гамма-системой, так как инициируется с мышечных рецепторов. Повышенная активность гамма-мотонейронов приводит к повышению возбудимости мышечных веретен и большей выраженности сухожильного рефлекса при механическом их раздражении молоточком. Изменение возбудимости гамма-мотонейронов сказывается на живости сухожильного рефлекса и не влияет на амплитуду Н-рефлекса. С учетом этой особенности при патологии возможно сочетание сохранного сухожильного рефлекса с отсутствием Н-рефлекса. Первично-мышечная патология с поражением мышечных рецепторов (воспалительного или дегенеративного характера) ведет к исчезновению сухожильного рефлекса при сохранном Н-рефлексе.
При патологии сегментарной дуги амплитуда Н-рефлекса снижается, а латентность возрастает. Снижение амплитуды Н-рефлекса связано с дисперсией возбуждающего разряда, идущего по сенсорным волокнам к мотонейронам (при полинейропатии). Неодновременность прихода активирующего импульса к мотонейронам ведет к недостаточно высокому постсинаптическому потенциалу, неспособному их активировать. Возникновение блока проведения по двигательному корешку S1 (реализующему Н-рефлекс в икроножных мышцах) также приводит к исчезновению Н-рефлекса. В этом случае и F-волна не будет регистрироваться, в то время как при поражении сенсорных волокон (S1 корешка) F-волна будет сохранна. Снижение амплитуды Н-рефлекса за счет нарушения проводящих свойств сегментарной дуги (как
интра-, так и экстраспинальной части) всегда будет сочетаться с увеличением латентности Н-рефлекса.
Снижение амплитуды может происходить и за счет центральных механизмов торможения, как правило, пресинаптического. В этих случаях латентность Н-рефлекса будет в пределах нормы.
Патологическое повышение амплитуды Н-рефлекса связано с повышением активности мотонейронов передних рогов спинного мозга за счет, как правило, снижения тормозных регулирующих влияний со стороны супрасегментарных структур. Патологическое повышение Н-рефлекса ведет к отсутствию депресии Н-рефлекса при низкочастотной (3 Гц) и супрамаксимальной стимуляции. Повышение амплитуды Н-рефлекса мышц голени сопровождается одновременным появлением Н-рефлекса в тех мышцах, где в норме он не регистрируется – мелких мышцах стоп и кистей. Система пресинаптического торможения мотонейронов нарушается в наибольшей степени на уровне спинальных мотонейронных пулов мелких мышц стопы, меньше – на уровне спинальных мотонейронных пулов мышц предплечья и еще меньше – на уровне пулов мышц кисти (М.Х.Старобинец и соавт., 1983).
Дата добавления: 2016-03-27; просмотров: 5681;