ХАРАКТЕРИСТИКА ВІРУСОПОДІБНИХ БІОПОЛІМЕРІВ ТА ВІРУСІВ 7 страница
Комбинация периодически повторяющихся саккадических и следящих движений называется нистагмом. При оптокинетическом нистагме, наблюдающемся при однонаправленном движении объектов в зрительном поле (например, при рассматривании растущих вдоль дороги деревьев из окна движущегося поезда) медленные следящие движения удерживают изображение объекта неподвижным на сетчатке, а быстрые саккады переводят глаза в исходную позицию. Другим видом нистагма является вестибулярный нистагм (см. стр. 238). Он вызывается раздражением вестибулярного аппарата, сопровождающим повороты головы, и возникает даже в совершенно темном помещении или при закрытых глазах. Этот вид нистагма выполняет ту же функцию удержания объекта в зоне
107
ясного видения при собственных движениях наблюдателя. На рис. 32 схематически изображены оба вида нистагма.
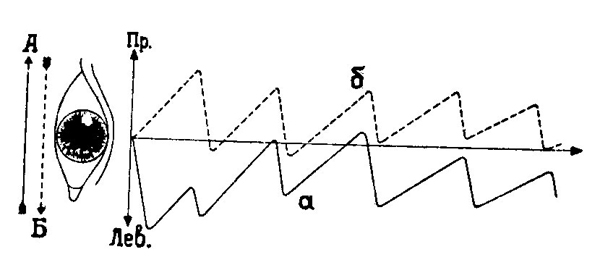
| Рис. 32. Оптокинетический (а) и вестибулярный (б) горизонтальные нистагмы (по Н. Бишофу, 1966). А — направление движения объектов, Б — направление движения головы. |
Вызываемый и регулируемый движением контуров по сетчатке оптокинетический нистагм отличается у человека от вестибулярного сильно выраженной тенденцией фиксировать появляющиеся на границе зрительного поля объекта. Поэтому нистагм осуществляется в этом случае на стороне быстрой фазы, которая также и начинает движение (рис. 33, а). Как показали исследования характеристики нистагма исключительно сильно зависят от расчлененности оптического поля, а также от степени внимания наблюдателя. Так например, отвлечение внимания для решения в уме арифметической задачи уменьшает амплитуду нистагма (Р. Кордс, 1926) или скорость следящих движений, так что саккадические движения могут практически исчезнуть (Г. Борриесс, 1923; С. С. Смирнов, 1971).
Второй класс движений глаз образуют микродвижения — мелкие непроизвольные движения глаз во время фиксации. Долгое время существование микродвижений оспаривалось, так как при фиксации неподвижной точки человек склонен считать свои глаза также неподвижными. Однако с развитием чувствительных объективных
108
методов регистрации движений глаз было доказано существование трех видов микродвижений глаз: тремора, дрейфа и фликов. Тремором называется дрожание глаза с частотой 20—150 гц и амплитудой порядка 10 угл/сек. Более выражены медленные дрейфы, со скоростью около 10 угл. мин/сек, и быстрые флики или микросаккады, амплитуда которых в среднем равна 7 угл. мин. Считается, что основная функция дрейфов состоит в дестабилизации ретинального изображения, микросаккады же возвращают его в зону наилучшей видимости. Комбинация дрейфов и фликов называется физиологическим нистагмом. По своим амплитудным характеристикам викарные движения занимают промежуточное положение между микро- и макродвижениями глаз. В их число входят движения глаз при стабилизации изображения относительно сетчатки, посттахистоскопические движения глаз, быстрые движения глаз во время сновидений и при галлюцинациях. С помощью викарных движений осуществляются преобразования информации, накопленной сенсорным звеном зрительной системы в виде следов раздражителя, а также манипулирование образам зрительной памяти.
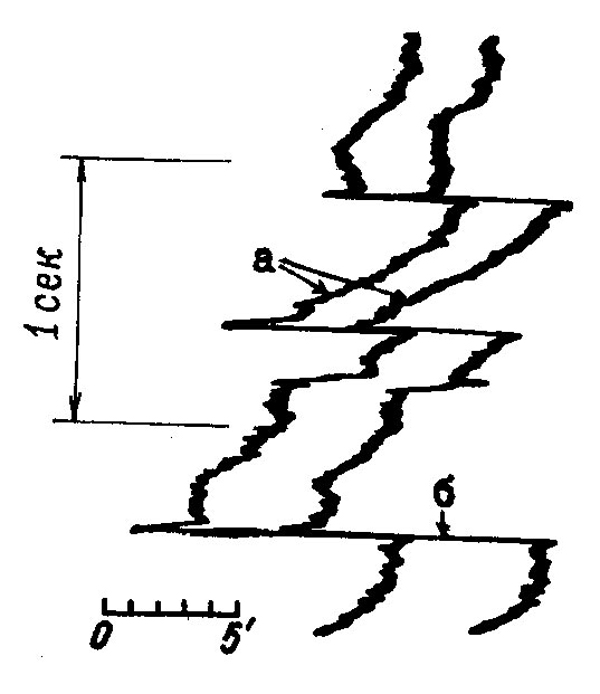
| Рис. 33. Физиологический нистагм: а — дрейф, б — микросаккады. Таким образом, движения глаз входят в число интимных механизмов зрения, обеспечивая как решение подсобных задач |
109
(например, аккомодацию, конвергенцию, дестабилизацию), так и процессы выделения существенных признаков воспринимаемого предмета и формирование активного зрительного образа.
Восприятие цвета
Зрительная система человека чувствительна к электромагнитным колебаниям, длина волны которых лежит в диапазоне от 380 до 720 нанометров (миллионных долей миллиметра). Эта область электромагнитных колебаний называется видимой частью спектра.
Рецепция падающего на сетчатку света представляет собой только первую ступень в сложной цепи процессов, приводящих к зрительному отражению окружающего нас мира. Структура процесса восприятия цвета меняется в зависимости от оптических свойств поверхностей предметов, которые должны быть восприняты наблюдателем. Эти поверхности могут светиться, излучая больше света, чем на них падает; блестеть, отражая весь падающий на них свет; отражать лишь часть падающего света и, наконец, быть прозрачными, т. е. не оказывать свету существенных препятствий.
Значительное большинство окружающих нас предметов относится к группе тел, частично поглощающих и частично отражающих падающий на них от искусственных или естественных источников свет. Цвет этих предметов объективно характеризуется их отражательной способностью. Поэтому для восприятия цвета предметов зрительная система должна учитывать не только свет, отраженный поверхностью предмета, но также характеристики освещающего эту поверхность света.
Однако в том случае, когда поверхность светится или, специально исключены признаки ее принадлежности какому-либо предмету, восприятие цвета может основываться лишь на анализе непосредственно излучаемого поверхностью света. Эта ситуация имеет место, если участок поверхности рассматривается через отверстие в большом темном или сером экране. Благодаря экрану скрадывается расстояние до поверхности и воспринимается диффузный цвет, относительно равномерно заполняющий отверстие. Такие цвета называются апертурными. Благодаря относительной
110
простоте процессов восприятия апертурных цветов, они изучены в настоящее время более полно, чем восприятие цвета предметов. Кроме того, исследование восприятия апертурных цветов имеет важное практическое значение, так как именно с ними приходится иметь дело операторам, работающим с современными индикаторами. Широкое использование цвета для кодирования информации связано с относительной легкостью различения апертурных цветов.
Цветовые ощущения, возникающие при восприятии апертурных цветов, полностью описываются тремя характеристиками или качествами. К ним относятся светлота, цветовой тон и насыщенность.
Первая из этих характеристик — светлота — иногда также не называется видимой яркостью. Светлота определяется прежде всего физической яркостью света. Как показали психофизические исследования, зрительная система способна реагировать на очень незначительные изменения яркости света: (дифференциальный порог яркости равен всего лишь 0,01). Как уже говорилось, с ростом физической яркости видимая яркость увеличивается медленно. Экспонента соответствующей степенной функции равна 0,33 (см. стр. 78).
На величину абсолютных порогов яркости решающее влияние оказывает состояние адаптации зрительной системы. Весь диапазон воспринимаемых яркостей от совершенно темного до слепяще яркого света благодаря адаптации охватывает огромную область от 10—4 до 106 нитх). На рис. 34 показано изменение нижнего и верхнего абсолютного порогов в зависимости от средней яркости фона, определяющей адаптацию.
Физиологические механизмы зрительной световой и темновой адаптации были рассмотрены ранее (см. стр. 83 и др.). Анализ динамики световой чувствительности при адаптации к темноте позволяет установить момент перехода от колбочкового к палочковому зрению. Для этого адаптированного к дневному свету наблюдателя помещают в полную темноту и периодически измеряют нижний
111
абсолютный порог яркости. Результаты измерений показывают, что вначале порог быстро падает, стабилизируясь на постоянном уровне через 8÷10 мин, а затем наступает вторичное резкое снижение порога, которое прекращается лишь через 30÷40 мин после начала адаптации (рис. 35).
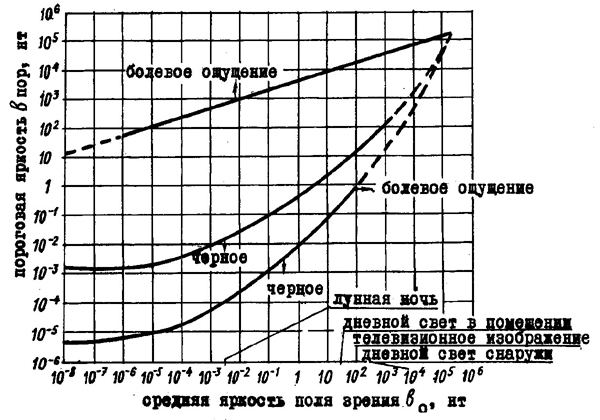
| Рис. 34. Границы зоны подлинных зрительных ощущений в зависимости от яркости зрительного поля, к которой адаптирован глаз (по Г. Н. Ильиной, 1972) |
Такой "двухступенчатый" вид кривая темновой адаптации имеет только тогда, когда пороги тестируются белым светом. Если используется красный свет, к которому палочки нечувствительны, кривая адаптации состоит только из своей первой ветви. Это доказывает, что точка перелома на кривой темновой адаптации соответствует моменту перехода от колбочкового к палочковому зрению.
Процесс световой адаптации продолжается обычно всего лишь доли секунды.
112
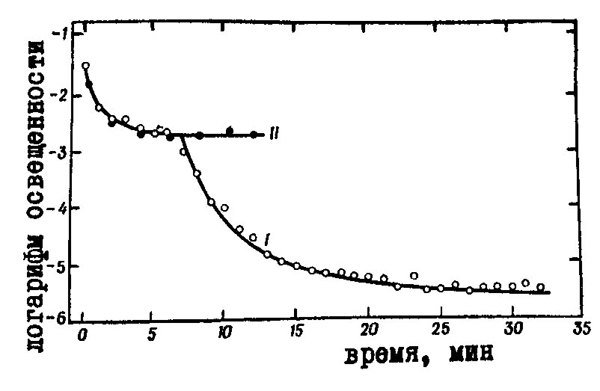
| Рис. 35. Изменение нижнего абсолютного порога яркости в ходе зрительной темновой адаптации: I — тестирование белым светом II — тестирование красным светом |
Видимая яркость меняется также в зависимости от длины волны раздражителя. При дневном освещении максимум спектральной чувствительности приходится на длину волны 555 нм, в темноте он сдвигается в коротковолновую часть видимого спектра к длине волны 510 нм (рис. 19). Это явление называется феноменом Пуркинье. Оно обсуждалось ранее в связи с многомерностью психофизических шкал (см. стр. 81 и д.).
Среди факторов, влияющих на светлоту, следует упомянуть размеры и время действия раздражителя. Эти факторы особенно важны для небольших и кратковременных раздражителей.
Влияние размеров выражается в увеличении вероятности обнаружения или видимой яркости пятна света с увеличением его площади. Этот эффект называется пространственной суммацией. Из него следует, что светлота раздражителя остается неизменной, если с уменьшением его яркости L одновременно определенным образом увеличивается его площадь S:
L·Sn=const
Показатель степени n, входящий в эту формулу, называется коэффициентом пространственной суммации. Область,
113
внутри которой n сохраняет свое наибольшее значение, равное единице, называется зоной полной пространственной суммации. В темноте при рассматривании источника света центральным зрением величина зоны полной суммации равна 15 угловым мин. Психофизиологические исследования показали, что величина зоны полной суммации определяется размерами рецептивных полей сетчатки. Поэтому она уменьшается в ходе световой адаптации и увеличивается при предъявлении раздражителя в периферическом зрении.
Аналогичная связь существует между яркостью и длительностью предъявления раздражителя:
L·Tm=const
Полная временная суммация (m=1) имеет место для экспозиций, продолжающихся 50—100 мсек.
Большой интерес представляют явления зрительного контраста. Одновременный или пространственный яркостный контраст заключается в подчеркивании зрительной системой различий яркости между соседними участками зрительного поля. Так, серый квадрат на черном фоне кажется светлее, чем такой же квадрат на светлом фоне (рис. 36).

Рис. 36. Яркостный контраст.
Американские исследователи Х. К. Хартлайн и Ф. Ратлиф при помощи электрофизиологических методов обнаружили наличие тормозных взаимодействий между рецепторными
114
элементами глаза пресноводного рачка limulus. Тормозное влияние, оказываемое рецептором А на рецептор Б, оказалось пропорциональным освещению А и пространственной близости обоих элементов. Это явление получило название латерального торможения (рис. 37).
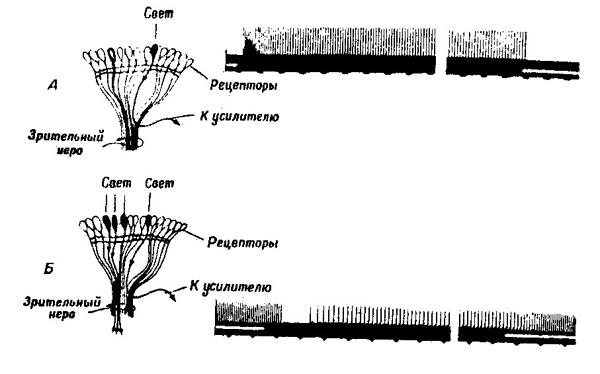
| Рис. 37. Латеральное торможение (по Ф. Ратлиффу, 1960). Освещение рецептора вызывает появление ритмических разрядов в соответствующем сенсорном волокне (А). Боковое освещение соседних рецепторов ведет к торможению ответа (Б). |
В результате латерального торможения, оказываемого соседними ярко освещенными элементами, расположенный на краю слабоосвещенной области рецептор будет разряжаться с меньшей частотой, чем элементы, освещенные столь же слабо, но расположенные дальше от границы двух областей. Напротив, рецептор, лежащий на краю ярко-освещенной области, будет разряжаться с большей частотой, чем рецепторы, расположенные в глубине этой области. Таким образом, благодаря латеральному торможению, картина возбуждений элементов сетчатки подчеркивает границы между областями различной яркости.
Тормозные взаимодействия были обнаружены в зрительных системах высокоорганизованных животных, включая обезьян. В то же время существуют данные о значительно более сложном, центральном происхождении явления
115
контраста.
Так, например, на контраст влияет сознательная установка наблюдателя. Если знак, изображенный на рис. 38, воспринимать как две латинские буквы V, то наблюдается выраженный яркостный контраст: левая буква кажется более светлой, чем правая. Если же воспринимать этот знак как одну букву W, то контраст исчезает.

Рис. 38. Влияние установки наблюдателя на яркостный контраст
(по К. Коффке, 1935).
Наряду с только что рассмотренным одновременным контрастом известен также последовательный яркостный контраст. Он выступает в виде последовательных образов — зрительных ощущений света, продолжающихся некоторое время после окончания действия раздражителя.
Различают отрицательные и положительные последовательные образы. Первые возникают, если при нормальном освещении в течение примерно 30 секунд рассматривать ярко освещенный объект, а затем быстро заменить его равномерным полем нейтрального цвета с более низкой яркостью. В этом случае испытуемый некоторое время видит перед собой темное пятно, по форме отдаленно напоминающее объект. Если же объект освещается в темноте вспышкой света, то возникает положительный последовательный образ. Как правило, он исчезает значительно
116
быстрее, чем отрицательный.
В последние годы нейрофизиологам удалось установить связь возникновения и смены последовательных образов с изменением активности нейронов в проекционных отделах зрительной коры (Р. Юнг, 1965).
Благодаря контрасту, зрительная система способна различать мельчайшие пространственные изменения яркости в зрительном поле. Способность видеть два близко расположенных объекта раздельными называется остротой зрения.
Острота зрения, при которой замечаются интервалы между объектами, величиной в одну угловую минуту, считается нормальной. Однако при благоприятных условиях острота зрения человека может достигать величины 0,5 угловой секунды. В этом случае расстояние между проекциями границ объектов на поверхности сетчатки в десятки раз меньше диаметра колбочки. Поэтому некоторые исследователи связывают столь высокую остроту зрения не с сенсорными процессами в сетчатке, а с микродвижениями глаз (см. стр. 107).
Острота зрения меняется вместе с состоянием адаптации. При низких уровнях освещенности она меньше, чем при высоких. На рис. 39 показано изменение остроты зрения для двух стандартных тест-объектов в зависимости от яркости фона. Особенно часто используется второй из этих тест-объектов — кольца Ландольта. Испытуемый должен указать положение разрыва на кольце.
Изменение остроты зрения в ходе адаптации соответствует изменению диаметра рецептивных полей.
Второй характеристикой апертуарных цветов является цветовой тон. Монохроматические, то есть вызванные светом, с одной длиной волны, красный, зеленый, желтый и другие цвета одинаковой видимости яркости различаются как раз по своему цветовому тону. Это качество цветовых ощущений связано прежде всего с длиной волны раздражителя.
При переходе от коротковолновых к длинноволновым электромагнитным колебаниям цветовой тон меняется следующий образом: раздражители с короткими длинами волн воспринимаются фиолетовыми, затем следует узкий участок чистого синего цвета, который кончается сине-зелеными тонами, далее находится узкая полоска
117
чистого зеленого цвета, за которым следуют желто-зеленые тона, потом появляется чистый желтый цвет и, наконец, в длинноволновой области — желто-красные цветовые тона.
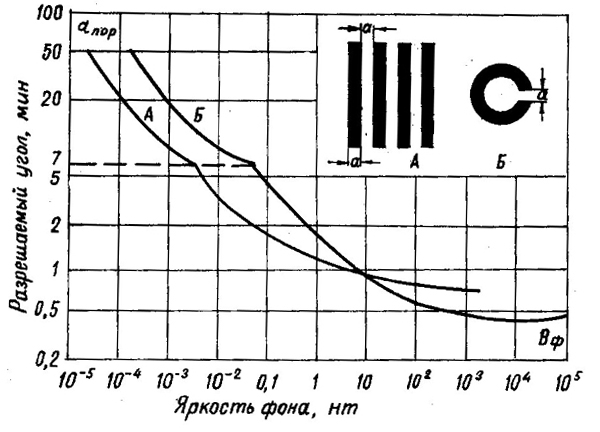
Рис. 39. Изменение остроты зрения в зависимости от яркости фона и типа тест-объектов
(по Г. Н. Ильиной, 1972).
Таким образом, монохроматические цветовые тона переходят друг в друга, образуя непрерывный ряд. Этот ряд можно превратить в замкнутый цветовой круг, если добавить к нему пурпурные (фиолетово-красные) цветовые тона, не являющиеся монохроматическими (рис. 40).
Зрительная система способна различать очень тонкие оттенки цветового тона. Общее число различных оттенков монохроматических тонов достигает 150—200. Минимальные разностные пороги, равные 1 нм, найдены в сине-зеленой (485 нм) и зеленовато-желтой (575 нм) частях спектра.
Если длина волны однозначно определяет цветовой тон, то обратное утверждение неверно. Одному и тому же цветовому тону соответствует бесчисленное множество различных комбинаций монохроматических раздражителей. Законы смешения цветов были открыты И. Ньютоном не позже 1692 года. Однако полностью их справедливость была доказана только в прошлом веке. Известны три таких закона:
118
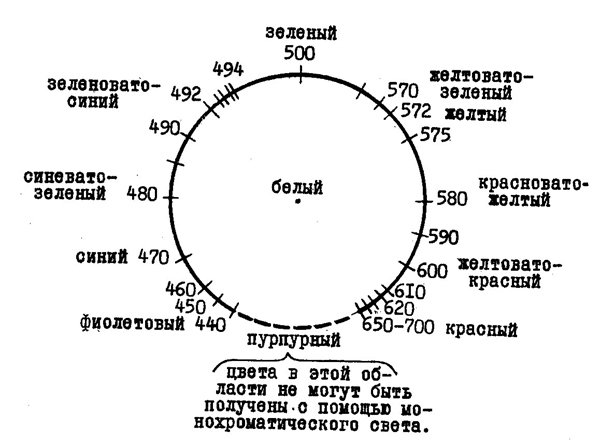
Рис. 40. Цветовой круг.
1. Для каждого цветового тона существует дополнительный цветовой тон, смешение с которым в определенной пропорции дает ощущение одного из оттенков серого (нейтрального) цвета. Следующие пары цветов являются дополнительными:
красный (660 нм) — сине-зеленый (497 нм)
оранжевый (610 нм) — зелено-синий (494 нм)
желтый (585 нм) — синий (485 нм)
желто-зеленый (570 нм) — фиолетовый (430 нм).
Легко видеть, что дополнительные цветовые тона расположены примерно на противоположных концах диаметров цветового круга.
2. При смешении двух цветов, лежащих на цветовом круге блике, чем дополнительные, цветовой тон смеси расположен между смешиваемыми цветами на соединяющей их прямой.
3. Одинаково выглядящие цвета, независимо от своего спектрального состава, дают при смешении одинаковые по цветовому тону смеси.
119
Наиболее важное следствие из законов смешения цветов заключается в том, что с помощью любых трех цветов, не являющихся дополнительными, можно получить любой цветовой тон. Тройки цветов, отвечающие этому требованию, называются основными цветами. К ним относятся, например, красный, синий и зеленый цвет. На рис. 41 показано, в каких пропорциях нужно брать монохроматические красный, зеленый и синий тона, чтобы получить все остальные цветовые тона видимого спектра.
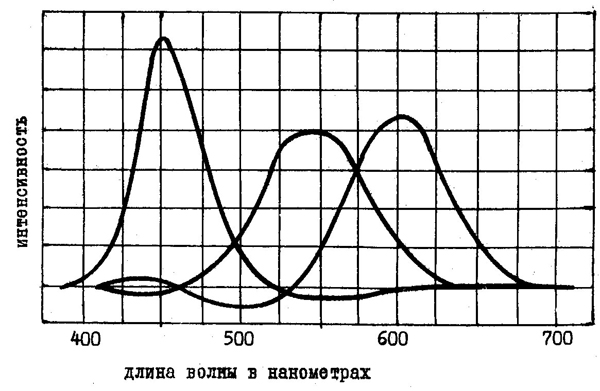
| Рис. 41. Пропорции красного (650 нм), зеленого (530 нм) и синего (460 нм) цветов, необходимые для получения всех цветовых тонов спектра. |
Степень отличия некоторого цветового тона от нейтрального тона, равного с ним по светлоте, определяет третью и последнюю характеристику цветовых тонов — их насыщенность. Физическим коррелятом насыщенности является "зашумленность" спектрального состава света электромагнитными колебаниями с другими длинами волн.
Насыщенность зависит также от яркости стимулов. Она максимальна для средних уровней освещенности и падает как при увеличении, так и при уменьшении яркости, вплоть до полного обесцвечивания
120
раздражителей. Синие, красные и пурпурные цвета кажутся сильно насыщенными и остаются насыщенными даже при низких уровнях яркости, желтые и зелено-желтые становятся относительно насыщенными при больших яркостях.
Как и в случае видимой яркости, существуют цветовая адаптация и цветовой контраст. Они выражаются в общем или локальном уменьшении воспринимаемой величины насыщенности цветового тона при его длительном рассматривании и в одновременном возрастании видимой насыщенности дополнительного цвета.
Цветовой последовательный контраст проявляется в виде цветовых последовательных образов. Например, длительная фиксация красного квадрата приводит к тому, что наблюдатель видит затем некоторое время перед собой зеленоватый квадрат, смещающийся вместе с движениями глаз. Еще И. В. Гёте обратил внимание на то, что цвета последовательных образов несколько отличаются от дополнительных, по сравнению с которыми они сдвинуты к краям спектра.
Все многообразие апертурных цветов, определяемых тремя рассмотренными характеристиками, можно представить в виде простой пространственной модели — цветового тела (рис. 42). Оно представляет собой двойную коническую пирамиду, по вертикальной оси которой происходит увеличение светлоты цвета. Каждое горизонтальное сечение цветового тела является цветовым кругом для данного уровня видимой яркости. Насыщенность цветового тона уменьшается при движении по радиусу цветового круга, в центре которого находится нейтральный серый цвет. Уменьшение радиусов цветовых кругов на концах цветового тела объясняется уменьшением насыщенности цветов при низких и высоких уровнях видимой яркости.
Вследствие того, что все цветовые тона, включая нейтральные, могут быть получены с помощью смешения трех основных цветов, на практике для описания цветов пользуются цветовым телом, сечение которого представляет собой не круг, а треугольник. На вершинах этого цветового треугольника находятся три основных цвета: красный, зеленый, синий.
На рис. 43 показан цветовой треугольник, принятый Международной осветительной комиссией (МОК) в качестве
121
эталонного. Условные коэффициенты на осях Х и Y определяют координаты каждого цвета внутри цветового треугольника. Укажем координаты цветности некоторых цветов:
| X | Y | |
| красный | 0,67 | 0,33 |
| зеленый | 0,21 | 0,71 |
| синий | 0,14 | 0,08 |
| голубой | 0,16 | 0,31 и т. д. |
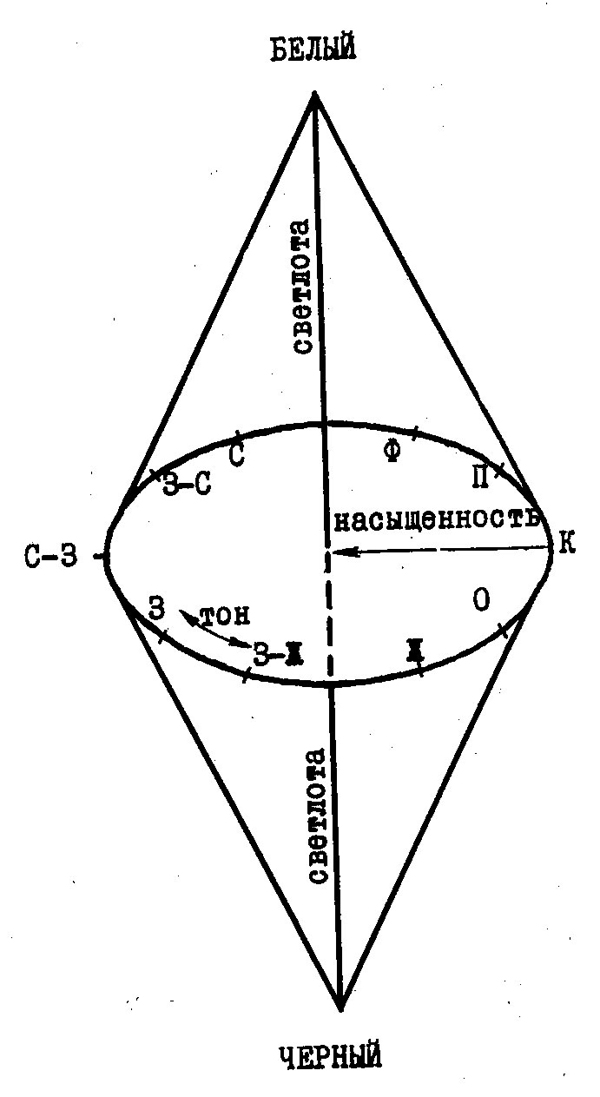
Рис. 42. Цветовое тело (см. текст).
122
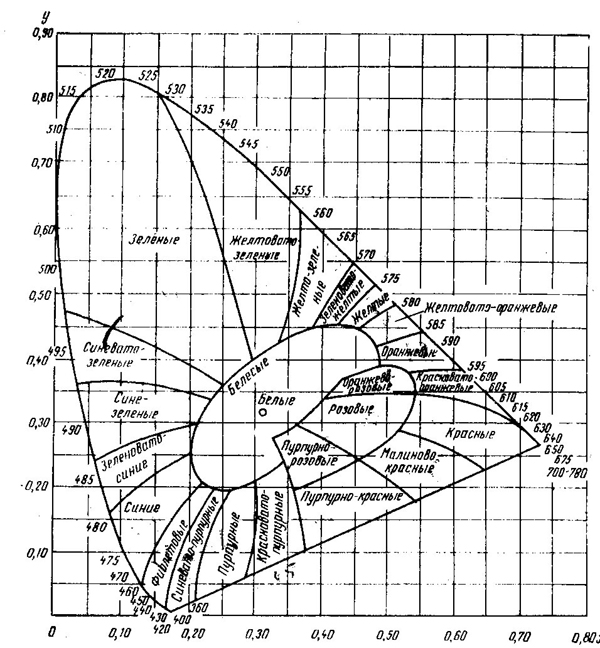
Рис. 43. Цветовой треугольник МОК (см. текст).
Существуют две классические теории цветовых ощущений, называемые трехкомпонентной теорией и теорией противоцветов.
Первые идеи о трехкомпонентности цветового зрения были высказаны М. В. Ломоносовым в его "Слове о происхождении света, новую теорию о цветах представляющем, июля 1 дня 1756 года, говоренном". Эта теория была детально разработана в XIX веке английским физиком Т. Юнгом и Г. Гельмгольцем.
Теория основана на предположении, что число различных рецепторов цвета в сетчатке должно быть невелико. Действительно, если предположить, что для каждого из воспринимаемых нами оттенков существует специальный рецептор, то в условиях монохроматического освещения работало бы меньше одного процента рецепторов и зрение должно было бы резко ухудшаться. Простые наблюдения показывают, что этого не происходит.
Так как все цвета могут быть получены с помощью смешения трех основных, то было сделано предположение, что в сетчатке
123
существуют три типа рецепторов, чувствительных к синему (фиолетовому), зеленому и красному цветам.
Альтернативную теорию выдвинул Э. Геринг (1878)х). В основу теории противоцветов легли данные о подробно изученных им явлениях контраста, а также некоторые психологические наблюдения. Так, большинство людей выделяют в качестве основного "главного" цвета, помимо красного, зеленого и синего цветов, также и желтый. Э. Геринг считал, что в сетчатке находятся три цветочувствительных субстанции. Их разложение приводит к восприятию белого, зеленого и желтого цветов, а восстановление — соответственно к восприятию черного, красного и синего цветов.
Обе теории долгое время противопоставляли друг другу. Одной из областей, в которой их сторонники искали подтверждения своих взглядов, было исследование различных аномалий цветового зрения.
Нарушения цветового зрения встречаются приблизительно у 8% мужчин и 0,5% женщин. Эти нарушения, по крайней мере отчасти, являются наследуемыми. Было бы неправильно называть этих людей цветослепыми, так как лишь один исключительно редкий вид расстройства цветового зрения связан с полной неспособностью различать цветовые тональности. Люди с такими недостатками называются монохроматами. В этом случае все длины волн и все смеси различаются исключительно по своей светлоте.
Значительная часть нарушений цветового зрения связана с затруднениями в дифференциации красного и зеленого цветов. Особые трудности эти люди испытывают при различении таких цветов, как голубой и розовый. Трехкомпонентная теория, исходящая из существования трех первичных видов рецепторов, объясняет эту аномалию выпадением рецепторов, чувствительных к красному или зеленому цветам. И действительно, было обнаружено, что существуют две разновидности красно-зеленой слепоты. В опытах на получение желтого цвета одним из этих цветоаномалов требовалось гораздо больше красного, а другим —
124
зеленого цвета, чем людям с нормальным зрением. Первая разновидность — нечувствительность к красному — была названа протанопией, а вторая — нечувствительность к зеленому — дейтеропией.
В пользу трехкомпонентной теории говорит, в свою очередь, существование слепоты на синий цвет, который путается в этом случае с зеленым. Это нарушение встречается почти столь же редко, как и полная цветослепота.
В то же время более детальные исследования показали, что красно-оранжево-желто-зеленая часть спектра преобразуется в восприятии цветоаномала не в оттенки зеленого (протанопия) или оттенки красного (дейтеропия), а в оттенки желтого цвета. Можно предположить, таким образом, что красно-зеленая слепота представляет собой дихроматическое желто-синее зрение. Этот факт в большей степени соответствует теории противоцветов Э. Геринга.
Аргументом в пользу теории противоцветов служат полученные на нормальных людях данные о порядке исчезновения цветового тона при перемещении стимулов в периферическое зрение. В этом случае первыми одновременно исчезают красные и зеленые цвета, от которых остается только желтый оттенок. Желтый и синий цветовые тона воспринимаются в более широкой области зрительного поля. Эти эффекты следует учитывать при использовании цветовой индикации.
Многочисленные доказательства как в пользу трехкомпонентной теории, так и в пользу теории противоцветов позволили Л. А. Орбели предположить, что справедливы обе теории. Однако каждая из них описывает закономерности переработки информации о цвете на различных уровнях зрительной системы. В последние годы детальное обоснование этой точки зрения было проведено американскими исследователями Л. М. Гурвичем и Д. Джемсон. На рис. 44 показана разработанная ими схема отношений между тремя светочувствительными субстанциями и четырьмя реципроктными процессами, лежащими в основе цветового зрения. Недавно были получены прямые нейрофизиологические доказательства справедливости этой модифицированной теории.
125
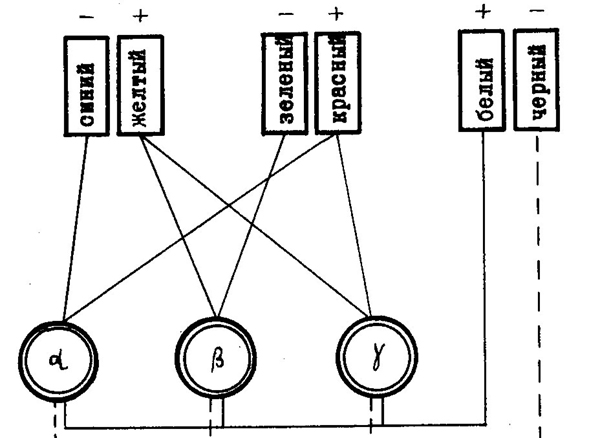
| Рис. 44. Упрощенная схема взаимоотношений между светочувствительными веществами и тремя парами противоположных процессов: сине-желтым, красно-зеленым и бело-черным (по Л. М. Гурвичу и Д. Джемсон, 1966). |
Прежде всего удалось показать, что в сетчатке действительно имеются три светочувствительных вещества. Один из самых тонких опытов в этой области был проведен английскими исследователями П. К. Брауном и Дж. Уолдом.
В их экспериментах миниатюрный пучок монохроматического света проецировался через зрачок на одиночные колбочки сетчатки испытуемого и с помощью микроспектрофотометра измерялось количество отраженного и вернувшегося через зрачок света. Было установлено, что существуют три типа колбочек, имеющих максимумы поглощения при 450, 525 и 535 нм.
Электрофизиологические опыты с микро-электродной регистрацией активности ганглиозных клеток сетчатки также говорят о существовании трех типов цветовых рецепторов. Шведский физиолог Р. Гранит показал, что возрастание активности нейронов возникает в ответ на освещение сетчатки синим, зеленым или красным светом (рис. 45).
126
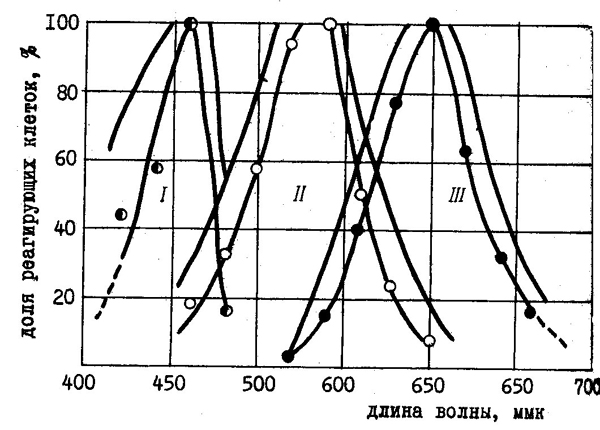
| Рис. 45. Регистрация импульсов одиночных ганглиозных клеток сетчатки (по Р. Граниту, 1955). После избирательной адаптации к соответствующим дополнительным цветам выявлено наличие рецепторов для синего (I), зеленого (II) и красного (III) цветов. |
Если исследования механизмов цветового зрения на уровне сетчатки подтверждают трехкомпонентную теорию, то исследования на более высоком уровне латерального коленчатого тела говорят в пользу теории противоцветов. Целый ряд работ, среди которых можно отметить исследования американского физиолога Р. Л. де Валуа и Е. Н. Соколова, показали, что на этом уровне наблюдаются реакции, оппонентного типа. Например, были найдены нейроны, увеличивающие активность в ответ на освещение сетчатки красным светом и уменьшающие ее в ответ на зеленый свет. Наряду с такими "красно-зелеными" элементами были найдены также "желто-синие" и "бело-черные" нейроны.
Дата добавления: 2015-03-19; просмотров: 1067;