ХАРАКТЕРИСТИКА ВІРУСОПОДІБНИХ БІОПОЛІМЕРІВ ТА ВІРУСІВ 6 страница
Как показали исследования, величина определяемого таким образом нейтрального раздражителя меняется в зависимости от того, какие раздражители сравниваются между собой. Если поиск производится среди тяжелых грузов, то и нейтральный раздражитель будет относительно тяжелым. Поэтому американский психолог Г. Хелсон (1947) назвал нейтральный раздражитель уровнем адаптации. Ему удалось показать, что уровень адаптации равен среднему геометрическому величин раздражителей, с которыми имеет дело наблюдатель. Добавляя к исходному набору раздражителей новые, большей или меньшей величины, можно получить закономерное повышение или снижение уровня адаптации.
Очень важную разновидность адаптационных эффектов открыл советский психолог Д. Н. Узнадзе (1931). В его экспериментах испытуемому от 10 до 20 раз предъявлялись в одних и тех же участках поля зрения два различных по величине объекта, например, большой и маленький квадрат. Затем неожиданно показывались два равных квадрата. Подавляющее большинство испытуемых сообщало в этом случае, что квадрат, предъявляемый на месте меньшего, кажется им больше. На основании этого и других подобных опытов можно сделать вывод, что перцептивные оценки в известной
87
степени определяются выработанным, на основании непосредственно предшествующих восприятий состоянием готовности воспринимать тот, а не иной объект. Это было названо Д. Н. Узнадзеустановкой.
Все виды изменения чувствительности, зависящие не от внешних условий, а от внутреннего состояния организма. Обычно эти изменения носят характер обострения чувствительности и поэтому их называют сенсибилизацией.
Можно выделить две основные формы сенсибилизации. Одна из них связана с действием физиологических факторов, а другая — с влиянием задач, стоящих перед субъектом и мобилизующих активность его восприятия.
К числу физиологических факторов изменения чувствительности относится прежде всего возраст субъекта. На ранних этапах онтогенеза она относительно низка. Процесс повышения чувствительности продолжается до 20—30 лет, после чего ее острота вновь снижается.
Вторым фактором, вызывающим стойкие изменения чувствительности, являются эндокринные сдвиги. Известно, например, что беременность обостряет вкусовую и обонятельную чувствительность и приводит к снижению зрительной и слуховой.
Исследования Б. М. Теплова и его сотрудников показали, что величина чувствительности является типологическим признаком, определяемым основными свойствами нервной системы. В частности оказалось, что у людей со слабым типом нервной системы чувствительность выше, чем у людей с сильным типом. Чувствительность также по-разному меняется в процессе утомления.
Экстренные сдвиги чувствительности можно получить с помощью фармакологических средств. Такие вещества как кофеин, фенамин, адреналин приводят к снижению абсолютных порогов. Введение других веществ, например, пилокарпина, повышает абсолютные пороги.
К сенсибилизации приводит также раздражение других органов чувств. Это явление, близкое к явлению синестезии (см. стр. 56), было подробно изучено советским физиологом С. В. Кравковым. Проведенные им исследования показали, в
88
частности, что звуковые раздражители повышают абсолютную и разностную чувствительность зрительного восприятия "холодных" — голубого и зеленого — цветовых тонов и снижает ее по отношению к "теплым" тонам — красному и желтому. Чем сильнее раздражитель, тем выраженнее этот эффект. В свою очередь слабые световые раздражители обостряют слуховую чувствительность. Известны факты повышения зрительной, слуховой, тактильной и обонятельной чувствительности под влиянием слабых болевых раздражителей.
Интересно, что сенсибилизация возникает даже под влиянием раздражителей, величина которых попадает в субсенсорный диапазон (см. стр. 69 и д). Так П. П. Лазарев обнаружил снижение абсолютных зрительных порогов под влиянием облучения кожи ультрафиолетовыми лучами.
Сенсибилизирующее влияние задачи, стоящей перед субъектом, связано с приобретением раздражителем определенного сигнального значения. В простейшей форме этот процесс выступает при условно-рефлекторном изменении чувствительности.
Такое условно-рефлекторное изменение чувствительности было, например, показано в опытах А. О. Долина, посвященных так называемым фотохимическим рефлексам.
Исходным для этих опытов был тот факт, что засвет глаза приводит к временному падению чувствительности. Однако, как показали опыты, если засвет глаза сочетать с каким-либо нейтральным раздражителем, например, звуком, то в дальнейшем изолированное предъявление звука может приводить к такому же изменению чувствительности, к какому раньше приводил засвет. А. О. Долину удалось также показать, что даже предъявление одного слова "свет" или близкого к нему по значению слова "пламя" может условно-рефлекторным путем изменить чувствительность глаза.
Обострение чувствительности к раздражителям, имеющим сигнальное значение, многократно отмечалось и другими исследователями. В опытах, проведенных Г. В. Гершуни и его сотрудниками, испытуемому предъявлялись два
89
относительно сильных световых раздражителя, между которыми иногда появлялась слабо освещенная точка. Освещенность этой точки была настолько незначительной, что она не воспринималась испытуемым. Однако, достаточно было начать сопровождать предъявление этой точки болевым раздражителем, чтобы она стала восприниматься.
В филогенезе жизненно важные раздражители, сигнализирующие о биотических факторах среды, воспринимаются организмом с наибольшей отчетливостью. Известно, что животные одного вида, но живущие в разных условиях, чувствительны к совершенно различным раздражителям. Так, например, домашняя утка чувствительна к растительный запахам, а кобчик, питающийся животной пищей, к гнилостным. С другой стороны, общий принцип ориентации в пространстве — эхолокация, обусловил развитие одинаково высокой чувствительности к ультразвуковым колебаниям у дельфинов и летучих мышей.
Развитие разнообразных форм чувствительности в онтогенезе человека также определяется требованиями действительности. В исследовании Т. В. Ендовицкой, проведенном в лаборатории А. В. Запорожца, было показано, что острота зрения зависит от места обнаружения сигнала в структуре игровой деятельности. В двух сериях этих опытов ребенок 5—6 лет должен был определить положение разрыва в кольце Ландольдта (см. стр. 116). Если в первой серии успешность ответа не имела никакого значения для игровой деятельности ребенка, то во второй серии местоположение разрыва было сигналом того, из какой дверцы выедет игрушечный автомобиль. Обнаружение разрыва становилось в этом случае перцептивным действием, включенным в игровую деятельность. Это приводило к значительному увеличению остроты зрения — так если в первой серии дети различали положение разрыва, когда кольцо было отодвинуто в среднем на 250—260 см, то во второй серии это расстояние увеличивалось до 300—310 см.
Не менее убедительны данные, свидетельствующие об изменениях восприятия, происходящих под влиянием задач, ставящихся теми или иными профессиями. Оператор радиолокационной станции, безошибочно определяющий направление и скорость одной из множества движущихся по экрану целей, или дегустатор, способный определить не только сорт винограда, из которого сделано вино,
90
но и место, в котором он вырос — все эти примеры показывают, что придавая значение отдельным сторонам раздражителя, выделяя характерные признаки ситуации, человек способен в значительной степени управлять своей чувствительностью.
91
ЗРИТЕЛЬНАЯ СИСТЕМА
Зрение позволяет человеку улавливать световые колебания и тем самым воспринимать предметы, расположенные на далеком расстоянии. Оно относится к числу наиболее важных видов дистантной рецепции. Зрительный аппарат развивается в филогенезе довольно поздно. Если у некоторых беспозвоночных его еще нет вовсе, а у отдельных позвоночных он уступает по значению обонятельному рецептору, то у высших млекопитающих (высшие обезьяны) и особенно у человека зрение начинает играть ведущую роль, и зрительное отражение предметного мира становится важнейшей формой отражения действительности.
Рассмотрим сначала строение аппаратов зрительной рецепции, чтобы затем перейти к детальному анализу основных законов функциональной организации зрительной системы.
1. Анатомо-физиологические основы зрения
Зрительный анализатор представляет собой сложную, включающую ряд последовательных уровней, систему физиологических механизмов. У наиболее примитивных организмов фоторецепторы распределены по всему телу или сосредоточены с обеих сторон переднего конца тела, образуя светочувствительные пластинки (рис. 21, а, б). Глаз человека имеет вид подвижного полого
Рис. 21. Этапы развития глаза.
92
шара, покрытого спереди прозрачной оболочкой — роговицей, и с небольшим зрачковым отверстием, через которое проникают световые лучи (рис. 22). Зрачок отделен от внутренней среды глаза прозрачной линзой — хрусталиком. Кривизна хрусталика изменяется с помощью внутренних мышц глаза, что обеспечивает разумное преломление света, давая возможность поступающим лучам точно доходить до выстилающей заднюю стенку глаза сетчатой оболочки.
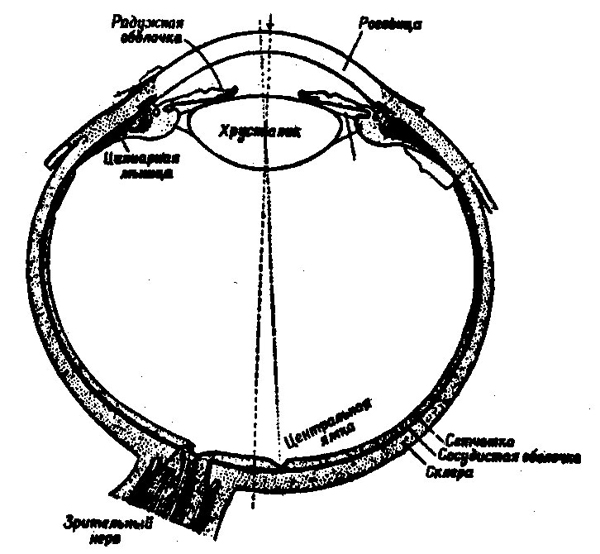
Рис. 22. Строение глаза
Сетчатая оболочка, или просто сетчатка глаза, представляет собой сложный нейронный аппарат, состоящий из нескольких слоев нервных клеток. Многие авторы с полным основанием расценивают ее как кусочек мозговой коры, вынесенной наружу (рис. 23).
Внешний слой сетчатки состоит из большого числа фоторецепторов, в состав которых входят специальные пигменты, например, зрительный пурпур. Разложение этих пигментов под действием фотонов света дает начало целой серии фотохимических реакций, приводящей к возникновению нервного возбуждения.
Различают два вида светочувствительных клеток: палочки (рецепторы ночного и сумеречного зрения) и колбочки (рецепторы дневного зрения).
93
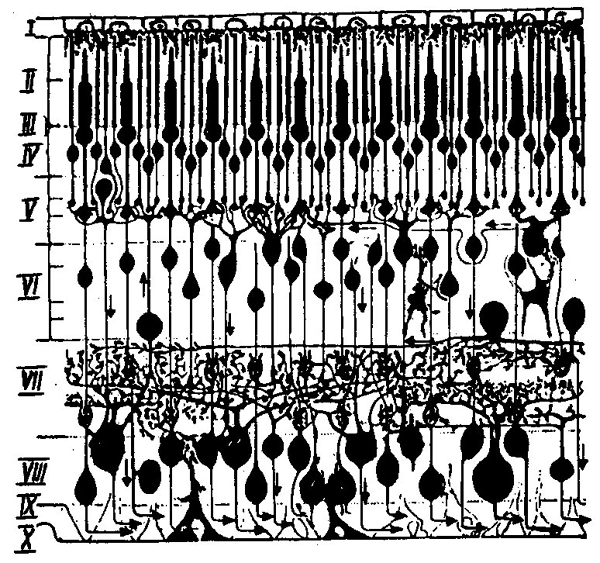
Рис. 23. Срез через сетчатку глаза.
В сетчатке находится около 130 миллионов палочек, которые рассеяны по всей ее поверхности. Они обладают высокой чувствительностью, т. к. содержащийся в них пигмент родопсин способен тонко реагировать на любые световые лучи, вызывая активацию сенсорных волокон. Колбочки находятся в сетчатке в гораздо меньшем количестве: их насчитывается до 7 миллионов. Они сосредоточены в центральных участках сетчатки, образуя область наилучшего видения или фовеа (рис. 24). Колбочки обладают относительно низкой чувствительностью, но зато содержащиеся в них пигменты могут избирательно реагировать на световые лучи различной длины, обеспечивая дневное или цветовое зрение. В силу того, что колбочки расположены лишь в центральной части сетчатки, мы можем различать цвета только в центральных участках поля зрения, на периферии зрительного поля удается различить лишь различные градации яркости, но не цвета объектов.
94
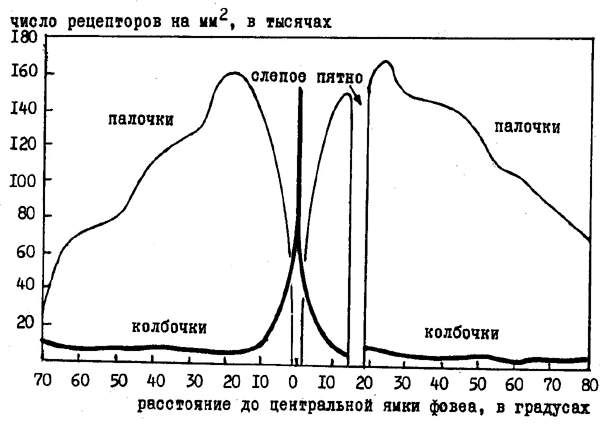
Рис. 24. Плотность распределения палочек и колбочек по поверхности сетчатки
Палочки и колбочки расположены в наружных слоях сетчатки (II—IV). Внутренние слои сетчатки (V—VIII) состоят из различного типа нейронов, по строению близких к тем, которые можно видеть в коре головного мозга. Они обеспечивают передачу и первичную обработку возникающего в рецепторах нервного возбуждения. Временную развертку соответствующих электрических процессов можно записать наложив один электрод на роговицу глаза, а другой установив на виске в виде ретинограммы (рис. 25).
95
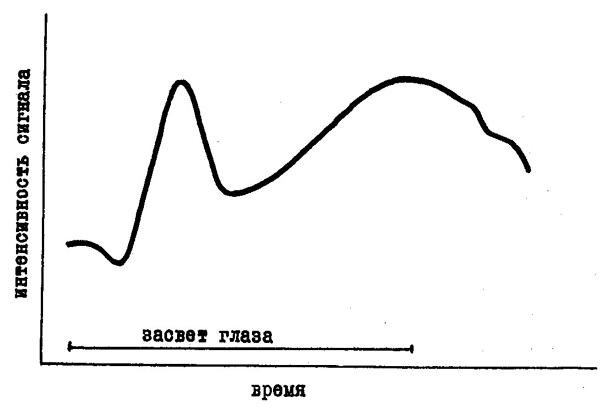
Рис. 25. Электроретинограмма человека
Нейроны внутренних слоев сетчатки выполняют различные функции. Одни из них, называемые биполярным клетками (VI), принимают и передают дальше возбуждения от групп соседних рецепторов. Количество рецепторов возбуждения, от которых суммируются на одном биполяре, или, как часто говорят, площадь рецептивного поля биполярной клетки зависит от их местоположения. В области фовеа каждая биполярная клетка обслуживает одну или всего лишь несколько колбочек. На периферии сетчатки на один биполяр приходятся десятки и сотни рецепторов. В отличие от биполяров, так называемые горизонтальные клетки (VI—VII), обладают мощным горизонтально расположенным дендритным аппаратом, позволяющим объединять или тормозить возбуждения, возникающие в разных группах рецепторов и биполярных клеток. Нейроны третьего типа — ганглиозные клетки (VIII) — не имеют непосредственных контактов с рецепторами. Они обеспечивают трансляцию зрительной информации в центральные отделы мозга.
96
Наконец, среди нервных клеток сетчатки имеются амакринные клетки (VI—VII), дендриты которых обращены к внутренним, а аксоны — к наружным слоям сетчатки. Благодаря этим особенностям строения амакринные клетки проводят возбуждения не от рецепторов к более высоким отделам нервного аппарата, а в обратном направлении. Поэтому они являются механизмами, позволяющими передавать установочные влияния центральных отделов нервной системы на светочувствительную периферию.
Приведенные данные дают основание для того, чтобы считать сетчатку глаза сложным рефлекторным прибором, обеспечивающим уже на периферии зрительной системы первичную переработку оптической информации.
Сетчатка глаза представляет собой периферический аппарат зрительной системы. Аксоны ганглиозных клеток, объединяясь образуют зрительный нерв. Он выходит из глаза недалеко от его центральной части. В месте выхода зрительного нерва сетчатка, естественно, не имеет светочувствительных клеток и поэтому оно представляет собой своеобразное "слепое пятно", не отвечающее на световые раздражители. Убедиться в этом нетрудно, если правым глазом фиксировать изображенный на рис. 26 крест. В какой-то момент можно заметить, что один из двух кругов сбоку от креста частично или полностью исчез: это означает, что его проекция попала на место выхода зрительного нерва — на "слепое пятно".

Рис. 26. "Слепое пятно" (см. текст).
97
После выхода из сетчатки глаза зрительный нерв проходит по пути, схематически изображенному на рис. 27. (Монокулярные поля зрения для удобства показаны на этом рисунке раздельно, хотя в действительности они накладываются друг на друга так, что их вертикальные оси совпадают). Сначала зрительные нервы обоих глаз идут порознь; затем они пересекаются. Это место называется перекрестом или хиазмой зрительных нервов. Характерным для строения этого перекреста является тот факт, что волокна, идущие от наружной части сетчатки каждого глаза идут в то же полушарие, а волокна от внутренней части сетчатки каждого глаза — в противоположное полушарие. Поэтому левая половина сетчатки обоих глаз (правая половина монокулярных полей зрения) представлена в левом, а правая половина сетчатки — в правом полушарии. В результате такого строения хиазмы и начинающегося от нее следующего этапа зрительного пути — зрительного тракта — поражения соответствующих разделов зрительного пути (опухолью, кровоизлиянием или травмой) приводят к своеобразным нарушениям зрения: поражение зрительного нерва вызывает слепоту на соответствующий глаз; поражение хиазмы приводит к выпадению обоих наружных полей зрения ("битемпоральная" или височная гимианопсиях)); поражение одного зрительного тракта — к одностороннему выпадению противоположной половины поля зрения. Эти симптомы имеют большое значение для диагностики топики (расположения) мозговых поражений.
Дальнейшие этапы центрального зрительного пути имеют сложное строение. Сначала зрительный тракт располагается на нижних (базальных) отделах лобной доли; затем он уходит вглубь мозга, отдавая часть волокон к среднему мозгу. Эти волокна заканчиваются в верхних буграх четверохолмия, являющихся элементарным рефлекторным центром зрительной системы. Этот центр играет ведущую роль у низших животных (например, у лягушки), а у человека сохраняет только простейшие функции, регулируя изменения диаметра зрачка и некоторые глазодвигательные рефлексы. Поэтому при поражениях верхнего четверохолмия зрачковый рефлекс выпадает. Основная масса волокон зрительного тракта направляется к подкорковым зрительным узлам: наружным коленчатым телам, в которых эти волокна пересекаются
98
и откуда начинается последняя часть зрительного пути — так называемое зрительное сияние. Оно веером проходит в глубине височных долей мозга и заканчивается в шпорной борозде затылочной области — первичном отделе зрительной коры.
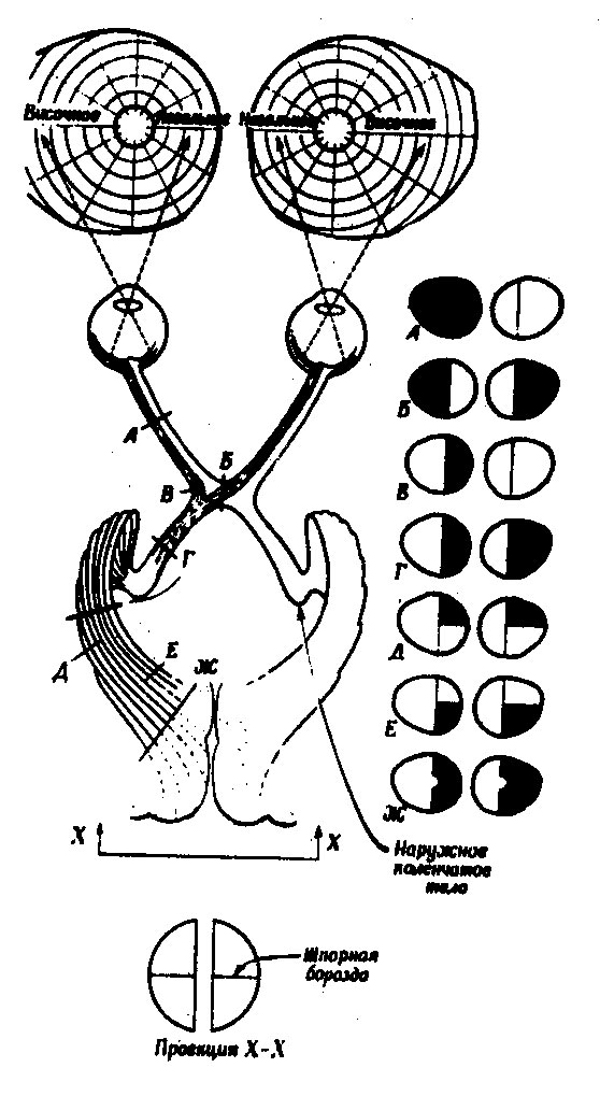
| Рис. 27. Схема зрительных проводящих путей, идущих к левому полушарию (по С. Дейчу, 1969). Справа темным выделены участки полей зрения, выпадающие при повреждении нервных путей в точках, указанных на схеме слева теми же буквами. А — полная слепота левого глаза; Б — двусторонняя височная гемианопсия; В — односторонняя назальная гемианопсия; Г — правая двусторонняя гемианопсия, возникающая в результате разрушения либо зрительного тракта, либо связей коленчатого тела со шпорной бороздой; Д и Е — гимианопсия правых верхнего и нижнего квадрантов; Ж — правая двусторонняя гемианопсия в результате повреждения обширной области в затылочной доле. |
99
Первичные отделы затылочной коры (поле 17-ое Бродмана) представляет собою конечный этап зрительного пути или, как его назвал И. П. Павлов, корковое ядро зрительного анализатора. Оно имеет своеобразное строение, отчетливо отличающее его от прилегающих отделов коры. В толще этого отдела коры особенно развит 4-й (афферентный) слой нервных клеток, в котором заканчиваются волокна зрительного пути. Как показали эксперименты американских физиологов Д. Х. Хьюбела и Т. Н. Визела, многие нейроны этой области отличаются высокой специфичностью: их рецептивные поля реагируют на такие частные признаки предметов, как округлые или острые линии, движение от центра к периферии или от периферии к центру и т. п. (рис. 28). Это дает зрительной коре возможность выделять из доходящей до нее информации огромное число составляющих ее сообщений.
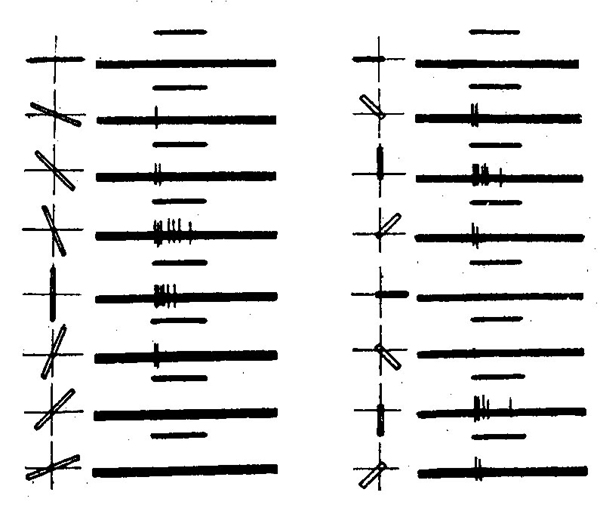
| Рис. 28. Детектор ориентации Хьюбела и Визела (по С. Оксу, 1969). Видно, что частота ответов нейрона максимальна при вертикальной ориентации полоски. Горизонтальное расположение не вызывает ответов, а промежуточное вызывает ответы меньшей интенсивности. |
100
Первичные отделы зрительной коры обладали и другим свойством, которое обычно обозначается как сомато-топичность: отдельные участки первичной зрительной коры поточечно представляют отдельные участки сетчатки. Поэтому поражение нижних отделов первичной зрительной коры вызывает выпадение верхних участков полей зрения, а поражение верхних — выпадение нижних отделов полей зрения.
Проекционные отделы затылочной коры являются лишь первым и наиболее простым этапом кортикальной переработки зрительной информации. Над ними надстроены вторичные отделы зрительной коры (поля 18-ое и 19-ое Бродмана). Отличительная особенность их строения заключается в том, что 4-й слой клеток здесь отсутствует и ведущее место начинает занимать 2-й и 3-й слой клеток, основную массу которых составляют клетки с короткими аксонами, которые несут объединяющую, ассоциативную функцию. Именно эти структуры и являются аппаратом, позволяющим объединять фрагментарную информацию, выделяемую клетками первичной зрительной коры, ставить ее под контроль более высоких инстанций мозга и комбинировать в динамические образы окружающих нас предметов.
Эта интегрирующая функция высших отделов зрительной коры с отчетливостью выступает из ряда физиологических и психологических опытов.
Как показал американский физиолог У. Маккаллок, кусочек бумажки, смоченный стрихнином и положенный на первичные отделы зрительной коры, вызывает возбуждение лишь в непосредственно прилегающих к нему тканях. Напротив, раздражение вторичных отделов зрительной коры вызывает или широко распространяющееся возбуждение, которое можно уловить на относительно далёком расстоянии (поле 18-е Бродмана) или торможение по всей прилегающей области (поле 19-е Бродмана)х).
Еще больший интерес для психологов представляют результаты психофизиологических опытов с раздражением соответствующих участков мозга.
101
Как показали наблюдения целого ряда неврологов и нейрохирургов (О. Пётцль, 1918, Х. Хофф, 1932, У. Пенфилд, 1950), раздражение первичных отделов зрительной коры, проведенное на операционном столе, вызывало у больных лишь неоформленные галлюцинации (светящиеся точки, шары, языки пламени), в то время как такое же раздражение вторичных отделов зрительной коры приводило к возникновению оформленных, иногда оценоподобных галлюцинаций (звери, люди, бабочки или целые сцены (рис. 29). Эти факты убедительно показывают, что первичные
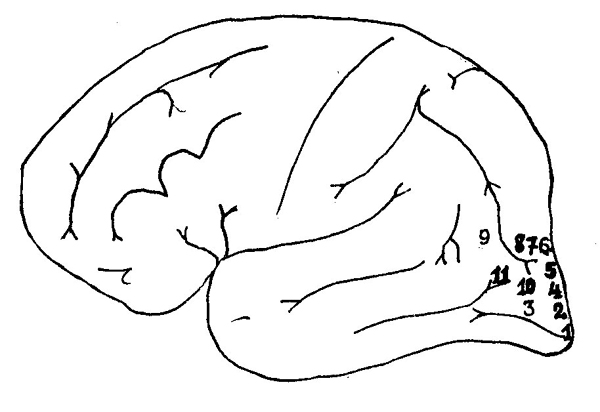
| Рис. 29. Схема точек затылочной коры, раздражение которых вызывает зрительные галлюцинации (по О. Пётцлю, Х. Хоффу и др.). Цифрами обозначены те участки мозговой коры, при раздражении которых возникали следующие зрительные галлюцинации: 1 — "Светящиеся шары". 2 — "Окрашенный свет". 3 — "Белый свет". 4 — "Голубой диск". 5 — "Пламя". 6 — "Голубой туман". 7 — "Лица, звуки голоса". 8 — "Идущий человек". 9 — "Лица". 10 — "Звери". 11 — "Лица и бабочки". |
102
и вторичные отделы зрительной коры по-разному участвуют в обеспечении зрительных перцептивных процессов, и что, если первичные отделы лишь выделяют доходящую до коры информацию, то вторичные отделы дают возможность превращать эти сведения в сложные структуры зрительного восприятия. Для изучения той роли, которую играют отдельные участки зрительной коры в процессе получения и переработки зрительной информации большое значение имеют данные нейропсихологических наблюдений, проводимых над больными с локальными поражениями этих зон мозга.
Как правило, поражение первичных отделов зрительной коры вызывает выпадение отдельных участков зрения, но никогда не приводит к распаду зрительного восприятия. Наоборот, поражение вторичных отделов зрительной коры, не вызывая частичных выпадений зрения, сопровождается распадом сложных форм зрительного восприятия. Больной с такими поражениями оказывается не в состоянии объединить воспринимаемые им фрагменты изображения в одно целое. Так, рассматривая рисунок, изображающий очки, он говорит: "кружок и еще кружок, перекладины..., наверное, велосипед?...", пытаясь угадать значение изображенного предмета там, где нормальный человек непосредственно воспринимает его. Эти факты приближают нас к пониманию мозговых механизмов зрительного восприятия. Они показывают, как происходит переработка зрительной информации и какие структуры мозга участвуют в этом сложном процессе.
Зрительное восприятие и типы движений глаз
Наблюдения показывают, что глаза человека никогда не остаются неподвижными. Как уже отмечалось, непрестанное движение является необходимым условием построения адекватного образа. Поэтому анализ зрительного восприятия невозможен без учета функционирования его моторного звена.
Подобно сенсорным механизмам зрительной системы, центры, управляющие движениями глаз, представлены на различных уровнях
103
ЦНС. Наиболее древние центры находятся в передних буграх четверохолмия. Они реализуют простейшие глазодвигательные рефлексы. Более высокий уровень глазодвигательного поведения, учитывающий характеристики объекта, возможен при участии глазодвигательных центров, связанных с затылочными долями коры мозга. Наконец, наиболее сложные — произвольные движения осуществляются при помощи структур передних глазодвигательных центров, расположенных в задне-лобных отделах мозга.
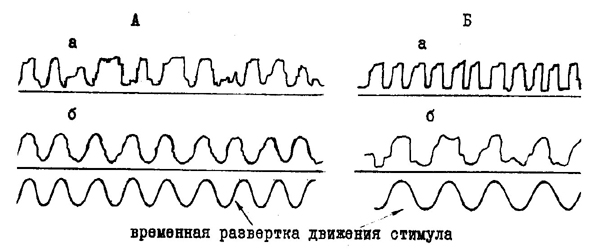
| Рис. 30. Движение глаза при произвольном переводе взора (а) и при отслеживании ритмически движущегося светового пятна (б) у больных с поражениями передних (А) и задних (Б) отделов мозга (по Е. Д. Хомской, 1969). |
Нейропсихологические исследования, проведенные советскими авторами, показали, что при поражении задних глазодвигательных центров существенно нарушаются пассивные движения глаз, следующие за движущимся предметом, в то время как произвольные движения глаз, совершаемые по речевому приказу (как можно быстрее переводить глаза из крайнего правого в крайнее левое положение и обратно) осуществляются значительно лучше. Наоборот, при поражении передних глазодвигательных центров следящие движения глаз сохраняются, а произвольное передвижение глаз отчетливо нарушается (рис. 30). Эти факты позволяют ближе подойти к важным мозговым
104
механизмам, регулирующим движения глаз и выделить основные факторы, лежащие в их основе.
Периферическую часть глазодвигательного аппарата образуют внутренние и внешние мышцы глаза. Первые находятся внутри глазного яблока. Из них цилиарная мышца изменяет кривизну хрусталика, обеспечивая резкость изображения на сетчатке при изменении расстояния до объекта, а мышца радужной оболочки глаза, меняя диаметр зрачка, регулирует общее количество света, поступающее на сетчатку.
Важную роль играют три пары внешних глазодвигательных мышц, содружественная работа которых приводит к изменению положения глаз в орбите. Выделяют три больших группы движений глаз. Первую группу составляют макродвижения глаз, впервые описанные И. Мюллером.
В число макродвижений прежде всего входят быстрые, скачкообразные саккадические движения. Они наблюдаются при зрительном поиске объекта, чтении или рассматривании неподвижных изображений (рис. 1 и 31). Амплитуда саккадических движений определяется размерами объекта и характером перцептивной задачи. Минимальная их величина равна долям градуса, максимальная — 40—60°. Обычно столь большие движения, возникающие как часть ориентировочной реакции на появление в периферическом зрении нового объекта, сопровождаются движениями головы и корпуса.
Латенный период саккадических движений равен приблизительно 150 мсек. Само движение осуществляется со скоростью от 50 до 100 град/сек. Характеристики саккадического движения задаются уже примерно за 50 мсек до его начала, так что если внутри этого временного интервала изменить положение цели, то глаза сначала прыгают на старое место, и лишь затем с обычным латентным периодом на новое место цели.
В последнее время (см. А. И. Назаров, 1971), было установлено, что незадолго до начала и во время скачка происходит повышение зрительных порогов. Этот феномен, названныйсаккадическим торможением, говорит о том, что прием зрительной информации осуществляется в промежутках между саккадическими движениями, когда глаза фиксируют ту или иную деталь объекта. Обычно глаза
105
находятся в режиме фиксации от 90 до 95% всего времени рассматривания объекта.
На рис. 31 показана запись движений глаз в процессе чтения. Видно, что глаз совершает серию фиксаций и скачков вдоль строки слева направо. По окончанию строки он производит большой скачок влево к началу следующей строки. Небольшое число возвратных движений может говорить о том, что читающий следит за смыслом текста и возвращается назад, чтобы выяснить непонятное.

| Рис. 31. Запись движений глаз в процессе чтения: а) на неподвижную, б) — на движущуюся фотобумагу. |
106
Когда объект, который необходимо рассмотреть, находится ближе и дальше от наблюдателя, чем точка фиксации, глаза поворачиваются так, чтобы новая фиксационная точка проецировалась на фовеальные участки. Этот вид макродвижений глаз называется вергентными движениями. Если точка новой фиксации находится ближе к наблюдателю, то движение глаз называетсяконвергенцией, если дальше — дивергенцией. Латентный период вергентных движений, как и саккадических, равен приблизительно 200 мсек, но их скорость относительно низка — от 5 до 30 град/сек. Вергентные движения возникают при удалении объекта от наблюдателя не более чем на 5—6 метров, на больших расстояниях оси обоих глаз практически параллельны.
Наконец, последним видом макродвижений глаз являются плавные и медленные следящие движения. Они возникают только в ответ на движение объектов или самого наблюдателя. В последнем случае следящие движения называются также компенсаторными, так как они противоположны по направлению движениям головы и способствуют сохранению фиксации объекта. Следящие движения возникают с латентным периодом равным 80—170 мсек и способны отслеживать объекты, движущиеся со скоростью до 40 град/сек. Во время слежения глаза могут отставать от объекта, особенно если скорость его движения велика. Возникающие движения изображения по сетчатке приводят к нарушениям различения тонких деталей объекта. Это явление, описанное американским психологом Э. Людвигом (1948), было названо динамической остротой зрения.
Дата добавления: 2015-03-19; просмотров: 1067;