Определение понятия и опыт классификации
Под совместным воздействием описанных в предыдущих главах факторов складывается тот или иной внешний облик растения, особенности ритма его жизнедеятельности, иначе говоря, присущая конкретному виду жизненная форма. Таким образом, жизненная форма растения может рассматриваться как результат адаптации ко всему комплексу экологических факторов. Один из лидеров науки о жизненных формах Е. Варминг (1884) трактует это понятие неоднозначно. Понятие "жизненная форма", по представлениям ученого - это то же, что и понятие "растительная форма", или "форма роста", и в то же время - это облик, габитуальная структура тех форм растений, которые в своей совокупности создают физиономичность растительных группировок. Е. Варминг по сравнению со своими предшественниками (А. Гумбольт, А. Гризенбах) существенно усилил эколого-физиологический аспект характеристики жизненных форм. В этой связи он акцентирует внимание на изучении приспособительных реакций растений к условиям среды. Именно в таком понимании жизненная форма стала объектом интенсивного и всестороннего исследования экологов, ботаников-географов, в том числе и фитоценологов.
Помимо общего габитуса, жизненная форма характеризуется физиологическими свойствами: листопадностыо, ритмом развития, продолжительностью жизни и др. Происхождение и родственные взаимоотношения в расчет не принимаются. Так, жизненная форма "листопадное дерево" объединяет деревья разных родов и семейств: березу, липу, платан, конский каштан и др. Общим, объединяющим признаком служит внешний вид, габитус - физиономия. Такое понимание жизненной формы давно и прочно вошло в практику и употребляется не только в специальной литературе, но и в учебниках.
Всестороннее и глубокое изучение растительного покрова и его компонентов изменило то содержание, которое первоначально включалось в понятие "форма растений" ("растительная форма"). Однако, как и прежде, это элементарная структурная, а затем и физиологическая (функциональная) единица растительного покрова. Вместе с тем растительная форма служит как бы показателем тех условий жизни, где она возникла и обычно распространена. С развитием представлений о биологической и экологической сущности единиц растительного покрова возникла необходимость более адекватной терминологии. Так, появилось понятие "экобиоморфа", под которым подразумевают типовую адаптационную организменную систему, сложившуюся в определенных условиях обитания.
Принципиально новое определение понятия жизненной формы дал тот же Е. Варминг в фундаментальной работе "Ойкологическая география" (1901). Совершенно справедливо он утверждал, что задача экологической географии состоит в выяснении формы приспособленности вида, т. е. его жизненной формы. Дальше, несомненно под влиянием работ Ч. Дарвина[*], он утверждал, что каждый растительный вид обладает определенными естественными, наследственными зачатками, вызывающими, благодаря каким-то внутренним, неизвестным нам причинам, образование форм, которые мы не можем поставить в связь с окружающими современными условиями жизни, а следовательно, и дать им научное объяснение.
На фоне всех предыдущих и большинства последующих определений становится особенно очевидно, что в данном случае Е. Варминг как бы совместил два понятия - "вид" и "жизненная форма". Причем последняя, по определениям ученого, выступает как элементарная единица процесса микроэволюции - вида, а не растительного покрова.
Итак, один из лидеров ботанической экологии Е. Варминг стал автором двух несовместимых определений понятия жизненной формы: первого - как единицы растительности (и это понятие все более совершенствуется в работах фитоценологов и экологов); второго - как элементарной единицы формообразовательного процесса (развивается исследователями микроэволюции, в том числе морфологами, изучающими филогенетический морфогенез, в частности трансформацию жизненных форм).
В науке, таким образом, сосуществует множество определений понятий "жизненная форма", или "биоморфа", но преимущественно в значении единицы растительного покрова. Приведем некоторые из них.
Известный советский ботаник-эколог Б. Келлер (1933) считал, что под жизненной формой нужно понимать определенную систему экологических приспособлений, тесно связанных с организационным типом растения, с принадлежностью его к определенному классу, семейству, а часто и роду.
С приведенной формулировкой полностью солидаризируется видный фитоценолог А. Шенников. Вместе с тем А. Шенников (1962) утверждает, что жизненные формы - систематические единицы в экологии, т. е. в экологической географии (а не в учении о виде). В. Алехин (1944), глава московской геоботанической школы, настаивает на адаптивной природе жизненных форм, считая, что это результат длительного приспособления растений к местным условиям существования, выраженный в его внешнем облике.
В разработку данной проблемы большой вклад внесли И. Серебряков и его школа. Основная концепция ученого изложена в фундаментальной монографии "Экологическая морфология растений" (1962). Обстоятельные формулировки структурной и функциональной сущности жизненных форм великолепно синтезируют те основополагающие принципы, которые выдвигали предшественники (Б. Келлер, В. Алехин, А. Шенников, а также О. Друде, X. Гаме, Г. Дю Рие и др.).
Солидаризируясь с А. Шенниковым в том, что жизненные формы - это систематические единицы в экологии, И. Серебряков взял на себя нелегкий труд создать всеобщую, четко дифференцированную систему жизненных форм. Система эта названа автором эколого-морфологической: "...она может быть полезной, - пишет И. Серебряков, - в ботанико-географических исследованиях и исследованиях фитоценологического характера". Определение понятия жизненной формы весьма обстоятельно. Под жизненной формой как основной таксономической единицей растительного покрова автор понимает совокупность взрослых особей данного вида в определенных условиях произрастания, обладающих, как уже было сформулировано выше, своеобразным общим обликом (габитусом), включая надземные и подземные органы (подземные побеги и корневую систему). Онтогенетически этот габитус возникает в результате роста и развития в данных условиях среды, а исторически - в определенных почвенно-климатических и ценотических условиях как выражение приспособленности растений к этим условиям.
Теперь вернемся ко второму определению жизненной формы, которое давал Е. Варминг (1901) под влиянием, как мы считаем, работ Ч. Дарвина. В этой формулировке, как отмечалось, речь идет о жизненной форме как элементарной единице микроэволюции - видообразования. Говоря о трудностях всестороннего исследования жизненных форм и вместе с тем перспективности выполнения этой задачи в будущем, он прямо выдвигает идею о том, что "...она же приводит к вопросу об исходных формах различных видов".
Такое понимание жизненной формы сложилось в сознании исследователей процесса микроэволюции (филогенетический морфогенез).
Таким образом, два взаимно несовместимых определения: первое - жизненная форма как основная таксономическая единица экологической системы растений, второе - единица микроэволюции видов процесса расообразования, в том числе эволюционного морфогенеза, - очевидны. Отметим, что исторически сложившееся противоречие, идущее от работ Е. Варминга, отразилось и в трактовке жизненных форм И. Серебряковым. Первая часть данного им определения традиционна и имеет в основном формальное значение. Вторая же часть оригинальная и представляет большой интерес. Становление жизненных форм понимается как этап развития (микроэволюции) в пределах популяционного арсенала биологического вида.
Следующий решающий шаг в формулировке понятия принадлежит В. Голубеву. Он приводит две трактовки понятия. Первая в эколого-биологическом (в узком смысле) - в традиционном понимании фитоценологов как биоморфы, возникающей конвергентно в пределах таксонов, не обязательно связанных родством. Изучение эволюции жизненных форм, понимаемых в этом смысле, интереса не представляет. Второе определение жизненной формы как категории морфобиологической в понимании исследователей формообразования и филогенетического морфогенеза таково: "в широком смысле жизненные формы представляют собою сложные системы морфофизиологических признаков, их совокупность, присущую в неповторимом разнообразии каждому виду и внутривидовым подразделениям. В характеристике этой системы находит отражение и уровень филогенетического развития данной группы растений, и их приспособление к условиям среды"; и далее: "изучение их эволюции неотделимо от анализа видообразования всеми доступными биологической науке методами". Более фундаментального определения жизненной формы научная литература не знает.
Приведенное определение В. Голубева основано не только на экспериментальных исследованиях онтогенетического и филогенетического морфогенеза - трансформации жизненных форм, но и на тщательном обобщении мирового опыта. Имеется в виду также опыт изучения формообразовательного процесса целой серии родов, внутриродовых систематических групп и отдельных полиморфных видов (линнеонов), представленных в разных участках ареала разными жизненными формами. В их числе и серия монографий, где эволюция жизненных форм от деревьев к травам представляется очевидным фактом. В сознании исследователей и комментаторов учения о жизненных формах изначально доминировало представление о формировании их облика под влиянием окружающей среды. Более того, высказывалось положение о том, что эта форма может эволюционировать в ходе онтогенеза особи. В этой связи следует также иметь в виду, что морфофизиологическое обособление жизненных форм происходит в определенных пределах под контролем генетического кода наследственности. Только с нарушением его функции, а также при сверхсильном химическом или лучевом воздействии наблюдается резкое, скачкообразное изменение на основе мутаций, рекомбинаций или полиплоидии.
И. Серебрякову принадлежит разработка иерархической таксономической системы жизненных форм, высшим таксоном которой, как и в систематике растений, служит отдел. Затем следуют соподчиненные таксоны: типы, классы, подклассы, группы, подгруппы, секции и, наконец, собственно жизненные формы. Таким образом, предложена дробная дифференциация биоморф до восьмой степени. Вместе с тем И. Серебряков - автор и наиболее детально разработанной классификации жизненных форм, в соответствии с которой выделяют четыре основных отдела: древесные растения, полудревесные, надземные травы, водные травы. Названные отделы охватывают восемь типов, в рамках которых осуществляется последовательная дифференциация биоморф.
В дидактических целях на уровне высшего биологического и сельскохозяйственного образования обычно предлагают более простую систему классификации жизненных форм датского ученого К. Раункиера. В основу его оригинальной классификации положен не внешний облик - габитус, а важные морфофизиологические особенности растений. Физиологический принцип предлагаемой системы - реакция растений на сезон покоя; морфологический - положение почек по отношению к поверхности почвы или верхушечных побегов в течение неблагоприятного для вегетации времени года. Система проста и обычно применяется на практике. Вместе с тем она наглядно демонстрирует позональную эволюцию жизненных форм покрытосеменных - от деревьев к травам. Микроскопические сравнительно-морфологические исследования И. Джеффри, затем Э. Синнота и И. Бейли подтвердили относительную древность древесных типов по сравнению с травянистыми. С другой стороны, состав жизненных форм может быть показателем климатических условий страны. Сам К. Раункиер рассматривал жизненные формы как феномен, возникший в ходе адаптивной (приспособительной) эволюции.
Согласно К. Раункиеру, жизненные формы растений можно подразделить на следующие пять главнейших типов: фанерофиты, хамефиты, гемикриптофиты и терофиты (от греч. phaneros - открытый, явный; chamai - низкий; hemi - полу; kryptos - скрытый; theros - лето). У фанерофитов почки переносят неблагоприятный период (мороз, засуха) "открыто", т. е. расположены достаточно высоко над почвой (выше 30 см). У большинства фанерофитов почки защищены специальными почечными чешуями, служащими для предохранения конуса нарастания и зачатков листьев от потери влаги.
Почки хамефитов располагаются почти на уровне почвы или не выше 20...30 см над ней. В холодном и умеренном климате эти почки часто получают дополнительную защиту в виде снежного покрова. Гемикриптофиты - травянистые многолетние растения, почки возобновления которых находятся на уровне почвы или погружены (неглубоко) в подстилку из растительного спада - еще одной дополнительной защиты. Криптофиты - травянистые многолетние и двулетние растения, почки возобновления которых полностью скрыты под поверхностью субстрата (почвы или воды). Терофиты - однолетние растения, переживающие неблагоприятный период в виде семян.
Классификация жизненных форм К. Раункиера в несколько сокращенном виде приведена на рисунке 21 и ниже.
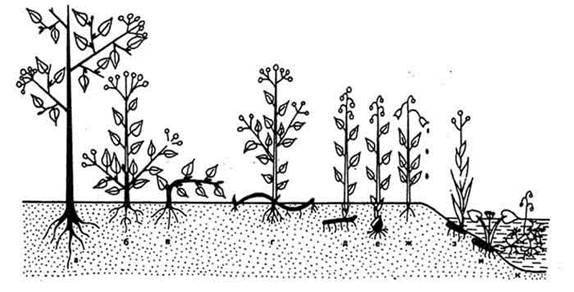
Рис. 21. Основные тепы жизненных форм, по К. Раункиеру: наземные растения: а - фанерофиты; б, в - хамефиты; г - гемикриптофиты; д, е -геофиты; ж - терофиты; прибрежноводные и водные растения: з - гелофиты; и, к - гидрофиты
Фанерофиты.
Мегафанерофиты (деревья выше 30 м высотой) и мезофанерофиты. (деревья от 8 до 30 м высотой).
1. Вечнозеленые, почки без почечных чешуй.
2. Вечнозеленые, почки с почечными чешуями.
3. Сбрасывающие листья, почки с почечными чешуями.
Микрофанерофиты (деревья и кустарники от 2 до 8 м высотой).
1. Вечнозеленые, почки без почечных чешуй.
2. Вечнозеленые, почки с почечными чешуями.
3. Сбрасывающие листья, почки с почечными чешуями.
Нанофанерофиты (кустарники менее 2 м высотой) и травянистые фанерофиты.
1. Вечнозеленые, почки без почечных чешуи.
2. Вечнозеленые, почки с почечными чешуями.
3. Сбрасывающие листья, почки с почечными чешуями.
4. Травянистые фанерофиты.
Эпифиты (от греч. epi - над), или растения, живущие на других растениях.
Хамефиты.
1. Кустарнички и полукустарнички; надземные побеги ортотропные, более или менее приподнимающиеся, не образуют подушек.
2. Пассивные хамефиты; надземные побеги ортотропные, но лежат на земле вследствие своей тяжести.
3. Активные хамефиты; надземные побеги плагиотропные, стелющиеся по земле.
4. Подушкообразные растения.
Гемикриптофиты.
1. Растения без листовых розеток:
o а) без плетей;
o б) с плетями.
2. Растения полурозеточные, с листовой розеткой внизу и с надземным облиственным побегом:
o а) без плетей;
o б) с плетями.
3. Растения розеточные, с листовой розеткой внизу, с безлистным или слабооблиственным надземным побегом, с симподиальным ветвлением.
4. Растения с надземными моноподиальными побегами и пластинчатыми листьями, но без чешуи:
o а) надземные побеги с листьями;
o б) надземные побеги без листьев (только с цветками):
§ без плетей;
§ с плетями.
5. Побег моноподиальный с пластинчатыми листьями и с чешуями:
o а) без плетей;
o б) с плетями.
6. Побег моноподиальный только с чешуями.
Криптофиты.
Геофиты (от греч. geo - земля) - почки возобновления находятся над землей.
1. Геофиты корневищные.
2. Геооиты клубнестеблевые.
3. Геофиты клубнекорневые.
4. Геооиты луковичные.
5. Геофиты с почками на корнях.
Гелофиты и гидрофиты (от греч. helos - болото, hydor - вода); почки находятся под водой.
1. Гелофиты - вегетативные побеги поднимаются над поверхностью воды.
2. Гидрофиты - вегетативные побеги погружены в воду.
Терофиты – однолетники, зимуют в стадии семян.
При изучении закономерностей распространения и распределения жизненных форм К. Раункиер разработал и применил метод статистического анализа. Построенные им биологические (фитоклиматические) спектры свидетельствуют о том, что в составе жизненных форм дождевых тропических лесов преобладают деревья (климат фанерофитов). Так, на Сейшельских о-вах на долю фанерофитов приходится 61 % общего числа учтенных видов, тогда как терофиты составляют лишь 16 %, а гемикриптофиты 12 %. В арктической зоне (о. Шпицберген) фанерофиты составляют 1 %, терофиты 2 %, а гемикриптофиты 60 %. Довольно широко представлены на Шпицбергене хамефиты - 22 % и криптофиты - 15 %. В умеренной зоне, например в Дании и России (Костромская обл.), преобладают гемикриптофиты - около 50 %, тоща как на долю фанерофитов приходится всего 7 %. В пустынях доминируют терофиты. Так, в Ливанской пустыне они составляют 42 % общего числа изученных видов (фанерофиты 12 %, хамефиты 21 %, гемикриптофиты 20 %, криптофиты 5 %).
Таким образом, в направлении от тропических дождевых лесов к арктической зоне наблюдается трансформация жизненных форм от мегафанерофитов к микрофанерофитам, нанофанерофитам и хамефитам, затем гемикриптофитам, геофитам и терофитам.
Дата добавления: 2017-02-20; просмотров: 587;