Происхождение и эволюция жизненных форм
В основополагающей работе "Происхождение видов" Ч. Дарвин стабильного термина для элементарных единиц формообразовательного процесса не дал. Биологические особенности такового он именовал по-разному: "зачинающийся вид", "форма", "сомнительная форма", "жизненная форма".
Теория о происхождении и развитии жизненных форм еще не создана. Вместе с тем давно уже на вооружении морфологов, изучающих жизненные формы растений, имеются достаточно основательные предпосылки для ее создания.
Отметим важнейшие из них. Теперь уже ни у кого нет сомнений, что с древних геологических времен до силурийского периода включительно длилось царство водорослей, среди которых древесных форм нет. Известно, что первые сухопутные растения - риниевые (Rhyniales) также были представлены своеобразными травами. Данными палеоботаники бесспорно доказано, что первые типично древовидные растения появились в среднем девоне, т. е. около 400 млн лет назад.
Первые древовидные были представлены высшими споровыми: плауновидными (Lycopodiophyta), хвощевидными, семенными папоротниками; следовательно, растительный покров состоял уже из многоярусных сообществ. Кроме первичных трав - риниевых, были распространены древесные и кустарниковые жизненные формы. В течение последующих геологических периодов состав всех ярусов существенно менялся за счет представителей новых систематических групп. Так, уже в каменноугольный период можно отметить бурный расцвет древесной растительности. Разнообразие древесной флоры резко возросло с появлением и развитием голосеменных: сосновых, саговниковых, беннетитовых (Benettttales), ганкговых, гнетовых (Gnetales) и др.
Важнейшие преобразования в составе жизненных форм произошли в мезозое параллельно с увеличением количества систематических групп голосеменных и широким распространением их представителей на континентах мира. Только в пределах порядков эфедровые (Ephedrales) и гнетовые отмечены следующие жизненные формы: деревья и деревца, лианы, кустарники и полукустарники. Однако травянистых форм среди голосеменных нет: ни первичных трав, ни вторичных.
Максимум многообразия современного спектра жизненных форм составляют вторичные травы. Они появились лишь со второй половины мелового периода на базе как высших споровых (травянистый вариант хвощевидных, плауновидных, папоротниковидных - Polypodyophyta), так и (особенно) покрытосеменных - победителей в жизненной борьбе.
Эволюцию жизненных форм в мире растений схематично можно представить в следующем виде:
· травянистые (водоросли, их производные - первичные наземные травы);
· деревянистые (высшие споровые и первичные семейства семенных);
· вторичные травы.
Главным объектом исследования жизненных форм служат биоморфы покрытосеменных, которые составляют неохватное многообразие и безраздельно господствуют в растительном покрове. Однако изучение эволюции указанной группы и путей ее исторического развития находится в неразрывной связи с не менее, а именно происхождением покрытосеменных.
Подавляющее большинство ученых-филогенистов - приверженцев противоборствующих гипотез согласны, что предками покрытосеменных обязательно были голосеменные. Имеются в виду, конечно, не ныне живущие, а давно вымершие их пращуры - первичные голосеменные. Постулируя данное положение, а также имея в виду, что среди голосеменных трав нет, легко прийти к заключению, касающемуся происхождения жизненных форм цветковых растений - покрытосеменных. Первыми обязательно должны были быть древесные жизненные формы, предположительно небольшие деревца.
Отсюда зародилась концепция об эволюции жизненных форм покрытосеменных от деревьев к травам. Упомянутая концепция гипотетически была высказана крупнейшими ботаниками-географами и интродукторами - Е. Джеффри (1899) и А. Красновым (1899) одновременно и независимо друг от друга. Первый утверждал, что с переходом от тропического климата к умеренному и холодному наблюдается "вырождение" древесных типов (исходных), что травянистые жизненные формы - новейшие, т. е. производные от древесных. Второй пришел к тому же выводу на основании исследования гистологических структур. Несколько позднее к аналогичному выводу пришел и крупнейший филогенетик Г. Галлир. Идея трансформации жизненных форм была подтверждена на разнообразных систематических группах покрытосеменных, преимущественно таксонах родового ранга. В результате М. Попов сформулировал стройную теорию ступенчатой трансформации жизненных форм покрытосеменных по схеме:

Следовательно, согласно М. Попову, в областях Старого Света развиваются две контрастные флоры по составу жизненных форм, видов и происхождению. Первая - жестко ксерофитная (жаро- и засухоустойчивая) в условиях ксеротермного климата пустынь и полупустынь ("флора вельвичии", см. с. 000). В плане теории эволюции жизненных форм вершиной аридной линии развития стали однолетние растения - эфемеры. Вторая - мезофитная флора ("флора гинкго") - развивается в более высоких широтах континента при относительно более влажных и менее жарких (лугово-лесных) условиях.
Последнюю линию развития М. Попов называл криофитной. Она хорошо прослеживается при экспансии покрытосеменных из стран умеренного (безморозного) климата в зимнеморозные климатические области. Трансформация жизненных форм в указанном направлении достигла высшего уровня в высоких широтах северного полушария по причине преобладания здесь суши и ярко выраженного резко континентального климата. В данном регионе нет однолетних трав, древесные растения представлены карликовыми формами, стланиками и подушками (рис. 22).
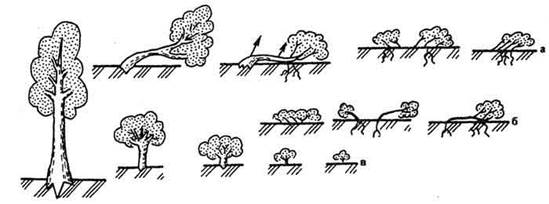
Рис. 22. Эволюционные изменения древесной формы роста в криофильных условиях: а, б - образование стлаников и подушек; в - нанизм (карликовость), по А. Хохрякову
Высказанное положение отмечали многие исследователи. В частности, Ф. Синнот и Дж. Бейли выяснили, что некоторые широко распространенные крупные по составу видов семейства в условиях северной умеренной зоны представлены целиком или почти целиком травами (табл. 1).
Таблица1
Состав (%) травянистых жизненных форм в некоторых семействах по регионам
| Семейство | Травянистые виды, % | |||
| Северная умеренная зона | Бразилия | Индия | Страны тропической Африки | |
| Астровые | ||||
| Молочайные | ||||
| Фиалковые (Violaceae) | ||||
| Бобовые | ||||
| Вербеновые (Verbenaceae) | ||||
| Вьюнковые (Convolvulaceae) |
Процесс редукции тела растений "от деревьев к травам" очевиден; он сопровождается также, как впервые отметил Н. Краснов (1889), уменьшением размеров репродуктивных органов, в частности цветка. Таков морфологический аспект теории эволюции биоморф (рис. 23).
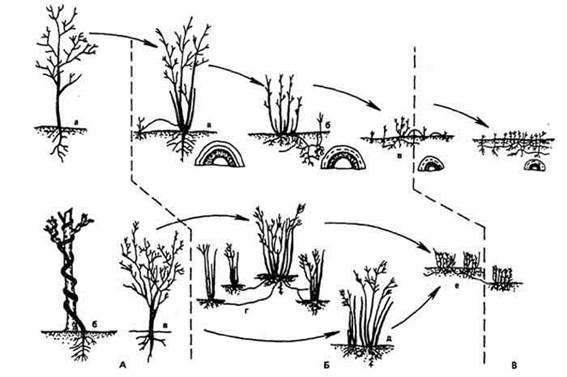
Рис. 23. Эволюция жизненных форм покрытосеменных на примере родов Rosa и Rubus:
А - мезофанерофиты, микрофанерофиты (вечнозеленые), Древнее Средиземноморье. Палеоген: а - Rubus arbor; б - Rosa moschata', в - шиповники из секции Canznae; Б - микрофанерофиты, нанофанерофиты, хамефиты (листопадные). Восточносредиземноморская, Иранская, Южноевропейская флористические провинции. Неоген: а - Rubus caucasicus; б - Rubus idaeus; в - Rubus saxatilis; г - Rosa cinnamomea; д - Rosa laxa; e - Rosa crenatula; В - гемикриптофиты, геофиты. Провинции: Степная, Таежная, Тихоокеанская. Антропоген: Rubus arcticus
Физиологический аспект сопряжен с изменением структуры растений, убыстрением темпов жизни, сокращением отдельных этапов развития и всего цикла от многих лет у вечнозеленых древесных форм до нескольких месяцев у однолетников, а в рекордных случаях - до трех-четырех недель у эфемеров. Генетический аспект - усложнение хромосомного аппарата, в частности дупликации - накопления копий одного гена, функционирование добавочных и рецессивных генов и др. Таков путь реального исторического развития - процесса, легко наблюдаемого в рамках подавляющего большинства древних таксонов, особенно у двудольных.
В этой связи напомним суждение К. Тимирязева, считавшего, что имеется два критерия совершенствования организмов: во-первых, усложнение организации, выражающееся в увеличении числа функций, в особенности в дифференцировке отправлений, а во-вторых, в лучшем приспособлений к условиям существования. Соматическая редукция (уменьшение и упрощение тела в процессе эволюции) у покрытосеменных в этом смысле, несомненно, прогрессивна. Она дает большое разнообразие обособленных форм жизни, более приспособленных к короткому лету и холодной зиме в высоких широтах континента (криофильная линия), равно как и в условиях короткого вегетационного периода в засушливых степных и пустынных районах Земли (аридная линия развития). На второй-третьей ступенях редукции от кустарниковых форм жизни к травянистым в связи с процессом геофилизации резко возрастает число органов (корневища, клубни, луковицы, выводковые почки), увеличивается число их функций, например разнообразие форм вегетативного размножения.
В практической деятельности земледельцев задолго до научных обобщений в области эволюции жизненных форм учитывалась закономерность развития - от деревьев к травам. Общеизвестны успехи в получении от деревьев и кустарников растений однолетней культуры. Таковы хлопчатник, клещевина (Ricinus communis). Изучение направления, темпов и причин трансформации жизненных форм имеет важнейшее практическое значение в селекционно-генетической работе. Учение о жизненных формах в рамках отдельных систематических групп, их организации, эволюции, многообразия типов на разнообразных экологических фонах, толерантности и реакции на всевозможные факторы среды составляет научную основу интродукции растений. Рациональное использование природных растительных ресурсов, например кормовых угодий - пастбищ и сенокосов, непременно связано с биологическим контролем за их состоянием, отражающемся в изменении видового состава, а следовательно, и спектра жизненных форм.
Результаты интересного исследования, косвенно подтвердившего теорию соматической редукции, опубликовал в 1956 г. шведский палеоботаник Хансен. Он разложил по спектрам Раункиера 38 000 видов растений ископаемой и современной флоры. Выяснилось, что самые молодые (четвертичный период) - это терофиты, геофиты и гамефиты, самые древние (мел) - мезо- и мегафанерофиты.
Ряд исследователей справедливо отмечают, что абсолютизировать трансформацию жизненных форм в рассмотренном направлении (от деревьев к травам) как единственно возможную было бы неверно, поскольку, во-первых, ряд крупных таксонов, охватывающих виды водных растений и эпифитов (нимфейные - Nymphaeaceae, орхидные, бромелиевые), не имеют, во всяком случае по современным данным, одревесневающих родичей (предков); во-вторых, известны случаи реверсии (от лат. reversio - возврат), когда из многолетних трав формируются кустарнички, например у некоторых бобовых в условиях аридного климата.
Контрольные вопросы. 1 .Какие принципы положены в основу классификаций жизненных форм растений? 2. Какие жизненные формы преобладают в умеренной зоне? В арктической зоне? В зоне пустынь? 3. Какие жизненные формы покрытосеменных эволюционно наиболее продвинуты? Примитивны? Какие доводы вы можете привести в доказательство правильности вашего суждения?
Дата добавления: 2017-02-20; просмотров: 3168;