Эфферентные связи прореальной извилины.
Нервные связи прореальной извилины с другими областями коры, а также с подкорковыми образованиями изучались многократно. После локального повреждения разных участков исследовалась дегенерация выходящих из нее эфферентных волокон.
Мы здесь приводим новейшие данные по результатам исследований А. Микеладзе и Г. Кикнадзе (34, 35) как эфферентных связей прореальной из-
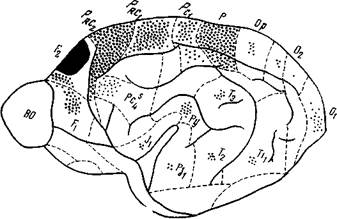
вилины (рис. 198 и 199), так и некоторых ее афферентных (рис. 200) связей. Эти авторы удаляли у собак часть прореальной извилины (поле F% по Адрианову и Меринг, 36) и прослеживали эфферентные пути методом дегенерации нервных волокон (по методу Наута в модификации Замбржицкого). Наблюдение за ходом и окончанием дегенерированных нервных волокон показало, что прореальная извилина посылает эфференты во многие корковые поля. Особенно тесная связь отмечена с полями, расположенными в непосредственной близости от удаленного участка коры. Это в первую очередь поле F^ (также расположенное в прореальной извилине), премоторное (prcz) и моторное (Рна) поля, поле Рс4 на орбитальной извилине и, наконец, теменное поле Р. В перечисленных участках коры дегенерированные волокна довольно многочисленны. Их основная часть расположена в VI—III слоях, меньшая же часть проходит через // слой и, не оканчиваясь здесь, достигает / слоя.
В более отдаленных от прореальной извилины полях (поля ОР, 01, 02 зрительного анализатора, поля сильвиевой извилины и др.) количество дегенерированных волокон резко уменьшается. В этих областях коры дегенерированные волокна достигают только нижних слоев коры (F—VI).
Обширные связи прореальной извилины с архипалеокортексом и подкорковыми образованиями известны по исследованиям многих авторов. Микеладзе и Кикнадзе (34) удаляли у кошек всю прореальную извилину, а у собак, как это было отмечено выше, только часаь извилины. На рис. 199 приведены результаты исследования этих авторов (обнаруженная ими дегенерация представлена в виде цепочек и точек) в сопоставлении с данными зарубежных авторов (дегенерированные участки даны стрелками).
|
Рис. 198. Эфферентные корковые связи прореальной извилины коры головного мозга собаки
Удаленный участок прореальной извилины (поле F2 по О. Адрианову и Т. Меринг) зачернен. Точками обозначены поля, где обнаружены дегенерированные нервные волокна, частота точек соответствует интенсивности перерождения волокон (Кикнадзе, Микеладзе)
Как видно на рис. 199, прореальная извилина у кошек и собак связана со многими подкорковыми образованиями. Густая масса перерожденных волокон направляется по внутренней капсуле. Отсюда часть их заворачивает в хвостатое ядро, а также в скорлупу и бледный шар. На этом уровне дегенерированные волокна найдены также в своде, прозрачной перегородке, передней комиссуре. Из внутренней капсулы перерожденные волокна достигают дорсомедиального и вентрального ядер зрительного бугра, они найдены в ретикулярном ядре зрительного бугра, но через это ядро они проходят не оканчиваясь, тогда как в первых двух ядрах волокна оканчиваются у тел нейронов. Кроме того, удаление прореальной извилины вызывает перерождение волокон в гипоталамической и субталамической областях, меньше в черной субстанции, а также у клеток красного ядра. Прореальная извилина связана также с собственными ядрами варолиева моста, с трапециевидным телом. Дегенерированные волокна найдены были в средней ножке мозжечка и его коре (подробно мозжечок не изучали). Многочисленная де-
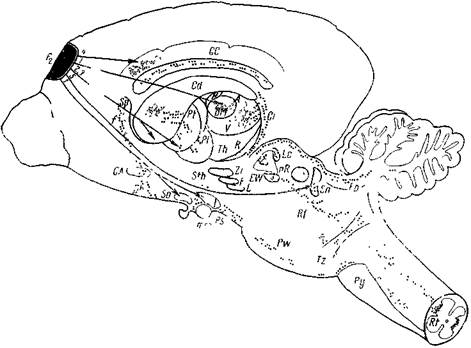
Рис. 199. Эфферентные связи прореальной извилины коры головного мозга кошин и собаки
Стрелками обозначены эфферентные связи прореальной извилины по данным литературы (/ — Аиег, 1956; 2 — Marshall, 1963; 3 — Webster, 1965; 4 — Cragg, 1965; 5 — Rinwik, 1966). Короткими цепочками обозначена проходящая, а мелкими точками — термин шьная дегенерация нервных волокон
| G С — поясная извилина; | L — Люисово тело; |
| Cd — хвостатое ядро; | ( — латеральный гипоталамус; |
| Pt — скорлупа; | So — супраоптическая область; |
| Р1 — бледный шар; | mm — сосковидные тела, |
| Sp — прозрачная перегородка, | Ps — ножки мозга; |
| С А — передняя комиссура; | Lc — синее вещество! |
| / — свод; | EW — ядра Здингер-Весфаля, |
| Db — диагональная связка; | nR — красное ядро; |
| Th — зрительный бугор; | Sn — черное вещество; |
| Н — уздечка; | Рр — ножка мозжечка; |
| А — переднее ядро зрительного бугра; | Л/ — ретикулярная субстанция, |
| DM — дорзомедиальное ядро зрительного бугра; | Pw — Варолиев мост; |
| R — ретикулярное ядро зрительного бугра; | Tz — трапециевидное тело; |
| Сг — внутренняя капсула; | Ру — пирамиды, |
| Sih — субталамическая область; | Rt — ретикулярный отросток серого вещества |
| ZI — неопределенная область | спинного мозга (Мике^дзе и Кикнадзе) |
| F — поля Фореля, |
генерация отмечается среди клеток ретикулярной формации, преимущественно в латеральных ядрах среднего и продолговатого мозга. Меньшая, но довольно четкая дегенерация прослеживается вдоль пирамид и в ретикулярном отростке серого вещества спинного мозга.
Микеладзе и сотр. (35, 37) исследовали также афферентную иннервацию прореальной извилины из разных участков коры и подкорковых образований. Результаты этих опытов подытожены на рис. 200. Хорошо видно, что с прореальной извилиной связаны не только ближайшая орбитальная извилина, но и сигмовидная извилина, сильвиевы извилины из новой коры, а из старой коры — поясная извилина и также такие подкорковые образования, как хвостатое ядро, субталамическая и заднегипоталамическая области, черная субстанция.
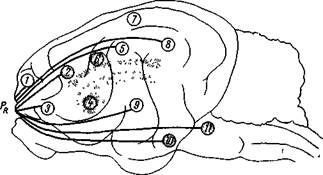
Рис. 200. Афферентные связи прореальной извилины коры головного мозга кошки
Кружочками обозначены исследованные участки: 1 — передняя сигмовидная извилина; 2 — передняя часть поясной извилины; з — орбитальная извилина; 4 — хвостатое 'ядро; 6 — передняя часть средней эктосильвиевой извилины 6 — средняя часть поясной извилины, 7 — средняя супрасильвиева извилина; 8 — задняя часть средней эктосильвиевой извилины; а — передняя сильвиева извилина; 10 — гипоталамическая и субталамическая области, 11 — черное вещество
Если принять во внимание, что прореальные извилины обширно связаны с премоторной и сенсомоторной областями коры, а также со всеми главными сомато-вегетативными координирующими механизмами подкорковых образований, то можно утверждать, что при активации всей объединенной системы субстратов памяти внешние реакции вызываются главным образом через прореальные извилины.
Надо думать, что через премоторные и сенсомоторные извилины наступают инструментальные движения конечностей и вообще локальные движения организма, а через кортикофугальные пути из прореальных извилин все сомато-вегетативные реакции эмоционального характера, а также такие общие для каждого поведения реакции, как локомоция, настораживание, ориентировочное движение головы и т. д.
На основании приведенных выше нарушений памяти по всем модальностям восприятия после удаления прореальной извилины и затем гистологических данных об афферентных и эфферентных волокнах прореальной извилины после локального повреждения коры мозга мы предполагаем, что все первичные зоны коры большого мозга двусторонне объединены ассоциационными нейронами не только со вторичными зонами, но и с нижневисочными извилинами и с прореалъными извилинами. Они должны возбуждаться совместно при каждом восприятии внешнего мира, и краткосрочная память, в основном, является совместной деятельностью этих корковых областей.
На рис. 196 была дана схема нервных субстратов коры и их нервных связей, в основном участвующих при зрительном восприятии объекта и репродукции его зрительного образа. И если принять во внимание обширные связи прореальной извилины с архипалеокортексом и подкорковыми образованиями, как это дано на рис. 199, то можно утверждать, что отсроченные реакции, возникающие на основе образной памяти, должно быть, происходят через эфферентную систему прореальной извилины.
Но фактически, как это будет видно ниже, воспринимающие области коры объединены и с корковыми отделами архипалеокортекса, (поясная извилина, гиппокамп), а также с подкорковыми образованиями (хвостатое ядро, амигдала и мозжечок). Двустороннее разрушение последних также приводит к временному исчезновению образной памяти. Но происходящие при этом нарушения образной памяти, должно быть, иного происхождения. Эти отделы старой и древней коры и подкорковых образований, должно быть, влияют на память тем, что они, действуя на новую кору своей афферентной импульсацией, поддерживают возбудимость нервных субстратов
образной памяти на некотором высоком уровне, необходимом для их актнвирования.
О роли гиппокампа в памяти.Вопрос о том, какую роль играет гиппокамп в процессе памяти, стал предметом специальных исследований после клинических наблюдений, когда у пациентов с пониженной функцией памяти отмечались дегенеративные изменения в гиппокампе. Изменения гиппокампа связывались особенно с нарушением памяти на недавние события. Они были отмечены и после таких нейрохирургических операций, когда по той или иной причине повреждался гиппокамп. Например, после удаления у пациентов энторинальной коры и гиппокампа сохранялась память на события далекого прошлого, тогда как выработка новых ассоциационных связей была очень сильно затруднена (Мильнер, 38).
Явные расстройства памяти на ближайшие события отмечаются в том случае, когда гиппокамп повреждается вместе с повреждением средней части височной коры(Сковилл и Мильнер, 39). Что при повреждении гиппокампа необходимо повреждение и других мозговых структур для нарушения памяти на это указывает следующий факт. При развитии амнезии в случае Корсаковского синдрома имеются дегенеративные изменения в гиппокампе; однако, как выяснилось, амнезия при Корсаковском синдроме наступает лишь в том случае, когда наряду с гиппокампом имеются изменения в таламических структурах (Виктор, 40).
Опыты на животных, пока еще недостаточные и не всесторонне проверенные, показывают неоднородную картину в изменении поведения после повреждения или удаления гиппокампа. У крыс после двустороннего повреждения гиппокампа отмечается некоторое затруднение в решении лабиринтной задачи —нахождения правильного пути (Каада и сотр., 41), однако такие крысы при выработке у них оборонительных условных рефлексов проявляют несколько лучшие способности как в процессе выработки этих рефлексов, так и в процессе восстановления угашенного рефлекса (Исааксон99, 100).
Возможно, что гиппокамп, хотя он принадлежит к структурам старой коры, приобретает наиболее важное значение в реализации памяти именно у человека. Здесь же следует отметить, что при повреждении гиппокампа фактически повреждается и перерождается множество путей, связывающих гиппокамп с разными структурами мозга. Поэтому при повреждении гиппокампа выключается ряд других образований.
Вопрос о роли гиппокампа в функции памяти требует дальнейших подробных и многосторонних исследований.
На основании всего вышеизложенного надо полагать, что все означенные нервные субстраты (вторичные зоны, прореальные извилины, нижневисочные доли и гиппокамповы извилины) объединены двусторонними связями и работают как единое функциональное образование в отношении осуществления образной памяти. Поэтому нарушение одного из этих звеньев ведет только к временному исчезновению или ослаблению образной памяти, ибо путем развития деятельности других звеньев происходит более или менее значительное восстановление.
Существование разных областей памяти в коре больших полушарий ставит вопрос — какая из этих областей является ведущей? Нам кажется, что по опытам на животных, на основании их пищевого поведения, мы можем говорить только о памяти на пищевые объекты. Есть основание предположить, что такой ведущий субстрат в отношении запоминания и воспоминания пищевого объекта и внешней реализации соответствующего поведения находится в префронтальной области, где первичной зоной вкусовых и тактильных восприятий из ротовой полости, а также двигательной зоной акта еды является орбитальная извилина. Прореальная же извилина, видимо, является вторичной зоной и в отношении этих вкусовых и тактильных восприятий. Она же, по-видимому, является вторичной зоной наряду с орбитальной извилиной в отношении обонятельной рецепции. У кошек в проре-
альной извилине также богато представлены большие пирамидные клетки, которые являются ассоциационными и характеризуют вторичные зоны (Каада, 42).
Известно, что орбитальная зона дает вторичные ответы на раздражение кожи (Аликишибекова, 43, и др.)) на звуки, на световые раздражения и на электрические раздражения зрительного нерва (Бюзер и Боренштейн, 44; Аликишибекова, 43, и др.), а на раздражение обонятельной луковицы реагирует вторичными ответами наряду с прореальной извилиной (Каада, 43). Наконец, мы сейчас знаем, что орбитальная и прореальная извилины производят синхронные медленные потенциалы в виде реакции вовлечения как спонтанно, так и на раздражение таламуса (Аликишибекова, 43).
Все приведенные факты свидетельствуют о том, что орбитальная и прореальная извилины работают как одна объединенная система, как это подобает первичной и вторичной зонам одной воспринимающей области, и в то же время здесь может происходить интеграция всех названных рецепций между собой и с эфферентным нейронным комплексом приема пищи.
Так как при каждом приеме пищи происходит возбуждение орбитальной и прореальной извилин зрительными, слуховыми, кожными, вкусовыми и обонятельными раздражениями пищевого объекта и одновременно возбуждаются соответствующие зрительные, слуховые, кожные, вкусовые и обонятельные воспринимающие области, то между всеми этими возбужденными отделами коры легко устанавливаются замкнутые нервные круги. И, очевидно, ассоциационные нейроны прореальной извилины, участвующие в этих кругах, испытывают вышеозначенное устойчивое изменение клеточных белков при первом же активировании данных кругов. Поэтому они могут обусловить репродукцию образа пищи и ее местонахождения в течение долгого времени и тем самым производят адекватные отсроченные реакции. За такое объяснение говорит уже описанный выше факт, что после двустороннего удаления прореальной извилины отсроченные реакции на зрительные, слуховые и вестибулярные восприятия пищевого объекта надолго исчезают.
Итак, судя по опытам на млекопитающих животных, первичные воспринимающие зоны совокупно со вторичными являются нервным субстратом восприятия и образов внешнего мира, а прореалъные извилины, должно быть, являются ведущим нервным субстратом памяти в отношении запоминания образа пищевого объекта и его местонахождения.
Дата добавления: 2017-01-13; просмотров: 1117;