О роли ассоциационных нейронов вторичных зон, нижневисочных долей и прореальной извилины.
Прямые опыты показывают, что не все ассоциационные нейроны имеют существенное значение в сохранении в памяти воспринятых объектов, т. е. в сохранении стойких активных белков в лостсинаптических участках. Такими ассоциационными нейронами прежде всего являются пирамидные нейроны, которые объединяют вторичные зоны воспринимающих областей с первичными.
Мы знаем, что у собаки после двустороннего разрушения вторичной зрительной зоны (18 и 19 полей) в течение первых послеоперационных недель краткосрочная память на зрительное восприятие местонахождения пищи исчезает. Она восстанавливается медленно в течение нескольких месяцев (Сихарулидзе, 30). Память восприятия другой модальности не нарушается. При этом животное видит, образуется условное пищевое поведение на зрительное раздражение. Существующие условные пищевые рефлексы не исчезают, но в первое время пищевое поведение по зрительной памяти исчезает полностью: животное идет к месту пищи, если видит. Но если спрятать пищу на его глазах, оно не пойдет туда спустя 10—15 сек. Лишь постепенно в течение 1, 2 или 3 месяцев краткосрочная зрительная память восстанавливается. Точно так же после двустороннего удаления вторичных слуховых зон (сильвиевой и супрасильвиевой извилин) временно исчезает краткосрочная память на слуховое восприятие, при этом память на восприятие другой модальности не страдает (Айвазашвили, 31).
После же двустороннего удаления префронтальной области, а именно ее прореальной извилины, сильно нарушается краткосрочная память на все виды восприятия: зрительное, слуховое, а также на вестибулярное, вкусовое и обонятельное, т. е. на комплексное восприятие пищевого объекта. Опять-таки это нарушение временное и через 2—3 месяца восстанавливается. И в этом случае условные рефлексы образуются и существующие не исчезают (Конорский и Лавицка, 32).
При удалении прореальной извилины, а затем вторичной слуховой зоны краткосрочная память страдает значительно сильнее в отношении слухового восприятия (Айвазашвили, 31).
Долгосрочная память, проявляемая после комплексного восприятия местонахождения пищи, в результате удаления вторичных зон или прореальной извилины по существу не нарушается. После комплексного восприятия пищи в совершенно новой обстановке такие оперированные собаки нойдут к месту пищи и в следующие дни.
Как указывалось выше, многократное повторение опыта с отсроченными реакциями на показ пищи в разнообразных ситуациях сильно повышает максимум отсрочки, т. е. краткосрочную память на зрительное восприятие нового места пищи. Поэтому можно думать, что после удаления вторичной зоны 18—19 полей деятельность прореальной извилины может развиваться в такой мере, что она одна сможет обеспечить репродукцию образа места пищи. То же самое должно происходить со вторичными зонами после удаления прореальной извилины.
В последнее время начали изучать образную память у собак после удаления из височных долей сильвиевой извилины и средней и задней частей эктосильвиевой извилины на обеих сторонах (Натишвили и Сихарулидзе, 33). Изучалась образная память как на изолированное слуховое, зрительное и вестибулярное восприятия, так и на комплексное восприятие со включением вкуса и обоняния. После операции животные жили 3—4 месяца и затем их забивали для морфологического исследования мозга.
У всех собак было выработано условнорефлекторное хождение к одной кормушке на определенный звук и к другой — на определенную фигуру
животного (кролика, кошки или собаки), проецируемую на экране. После операции наблюдалось сильное расстройство поведения. Они были неподвижны, инертны и боязливы. Некоторые стали агрессивными. Кроме того, все они казались совершенно слепыми. Через 2—3 недели после операх ии они стали видеть только на очень близком расстоянии — на 50—80 см. Через3—4мемяца они уже хорошо видели на расстоянии 2—3 м, но на большем расстоянии не могли видеть. Через месяц после операции поведение их изменилось к лучшему: они свободно двигались по экспериментальной комнате. Страх и агрессивные реакции исчезли. На окрик заходили в клетку. На условные сигналы также стали реагировать побежкой к соответствующей кормушке. Но тонкая дифференциация звуковых сигналов, а также дифференциация фигур не восстановились и через 3—4 месяца.
Одновременно сильно была нарушена образная память. В течение почти одного месяца на условные сигналы животные выходили из клетки только во время сигналов или в продолжение 10—15 сек. после них. Максимум отсроченной реакции на условные сигналы увеличивался постепенно. Спустя 3—4 месяца на условный звук собаки выходили и шли к кормушке только через 3—4 мин., в то время как до операции они выходили и через 10—12 мин.
При изучении краткосрочной памяти на зрительное восприятие местонахождения пищи максимум острочки был 6 мин. До операции они шли и через 25 мин. На постукивание миски собаки шли через 3—4 мин., в то время как до операции они шли через 10—12 мин. Максимум отсрочки на лабиринтную рецепцию в норме был 5—6 мин., а через 4 месяца после операции—2—3 мин.
Краткосрочная память на комплексное раздражение также нарушалась. Оперированная собака направлялась к месту пищи в течение 9 — 10 мин., в норме же так поступали и через 40 мин.
Но что поразительно, долгосрочная память у оперированных собак фактически не нарушалась. Они правильно направлялись к месту пищи на другой и на третий день. Это наблюдалось и через две-три недели после операции, когда краткосрочная память ограничивалась несколькими секундами (Натишвили и Сихарулидзе, 33).
Итак, двустороннее повреждение нижних отделов височных долей приводит к значительному ухудшению краткосрочной памяти без повреждения долгосрочной.
Итак, ассоциационные нейроны вторичных зон, прореалъной извилины и нижневисочных долей должны служить основной составной частью образуемых между этими участками коры замкнутых нейронных кругов. Долговременное вращение возбуждения и вызванная им длительная синаптическая потенциация с длительным повышением возбудимости в данных ассоциационных нейронах должны обусловливать краткосрочную память.
На рис. 196 дается приблизительная схема нервных элементов, участвующих в происхождении образной памяти на зрительное восприятие.
В замкнутых нейронных кругах,, образуемых при восприятии местонахождения пищи нервными субстратами памяти, ни проекционные пирамидные нейроны коры, производящие ориентировочные реакции (например, к летки Мейнерта и Кахала), оборонительные и инструментальные движения конечностей, а также другие внешние реакции, ни сенсорные нейроны с околоклеточной аксонной сетью, производящие восприятие, прямо не должны участвовать. Все эти нервные круги всецело состоят из вставочных и ассоциационных пирамидных нейронов. Но эти нейроны связаны своими коллатералями непосредственно или через добавочные вставочные нейроны с проекционными и сенсорными нейронами. Поэтому распространяющееся по нейронным кругам возбуждение в то же время активирует как сенсорные, так и проекционные нейроны. Сообразно этому одновременно репродуцируется соответствующий образ и наступает адекватная ориентировочная реакция.
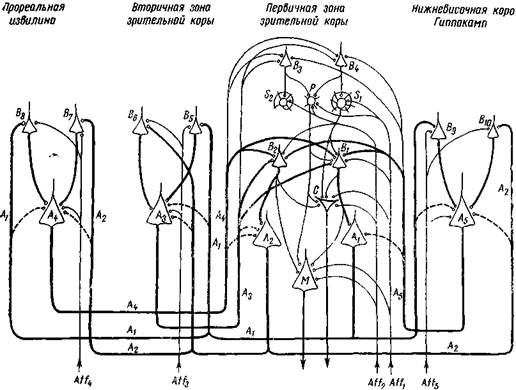
Рис. 196. Схема нервной структуры зрительной памяти
Участвуют первичная зрительная зона, вторичная зрительная зона, прореальная извилина, нижневисочная доля и гиппокамп.
б! и S2 — сенсорные звездчатые нейроны;
Р — передаточный звездчатый нейрон, В, — Вц, — вставочные пирамидные нейроны; ai — At — ассоциационные пирамидные нейроны;
С — проекционные звездчатые нейроны Кахала для движения глаз; М — проекционный пирамидный нейрон Мейнерта для движения шеи при ориентировочной
реакции;
a//j и A//z — афференты зрительной специфической системы; a//s, Aff,, Affs; — афференты неспецифической системы
Толстыми линиями обозначены волокна и клетки нервных кругов, активируемых при зрительном восприятии или при зрительном образе. Тонкими линиями обозначены односторонние связи нервных кругов с звездчатыми нейронами, производящие образ, и с проекционными нейронами ориентировочной реакции. Штриховыми линиями обозначены возможные коллатеральные связи замкнутых нейронных кругов.
Системы ассоциационных клеток А,, А4, иА5, должно быть, служат для удержания в памяти зрительных образов. Воспроизведение их при активировании одной из частей объекта происходит следующим образом: например, при воздействии части объекта через Affi сначала возбуждаются первичные зоны Si, Р, В], С, М и а!, возникает восприятие этой части и вызывается (через С и М) адекватная ориентировочная реакция Затем возбуждение переходит через А» во вторичную зону, активируя В, и А3, в прореальную извилину, активируя В, и А,, и еще в височную долю — гиппокамп — В, и А8. Из А3, А» и А5 импульсы идут обратно в первичную зону и активируют, во-первых, Вг и В» и втим путем приводят в активное состояние всю систему нейронных кругов и, во-вторых, через В, и Bt активируют системы звездчатых клеток Si, S2 и Р, репродуцируя образ всего объекта и вызывая адеьватную ориентировочную реакцию
Дата добавления: 2017-01-13; просмотров: 650;