Формирование в ходе изучения дисциплины "зоология беспозвоночных" социально-личностных компетенций 17 страница
Суть второй группы гипотез сводится к тому, что турбеллярии произошли от предка, имевшего целом и кровеносную систему в результате вторичного упрощения. Сторонники этих гипотез полагают, что главным фактором, обусловившим подобную трансформацию, могло стать уменьшение размеров тела, которое само по себе могло привести к утрате целомической полости и утрате транспортных систем (кроме диффузии). Турбеллярии, как предполагается этими гипотезами, взяли начало не от взрослых целомических животных, а от их личинок, то есть в становлении турбеллярий имела место неотения. Личинки целомических животных обладают малыми размерами, у них, как правило, сильно развиты ресничные образования, а целом и транспортные системы до поры до времени не развиты. Предполагается, что именно эти особенности унаследовали турбеллярии. Тем не менее, вдумчивый слушатель уже отметил, что в этой гипотезе не объясняются причины заднего положения рта и отсутствия пищеварительной полости.
В заключение считаю целесообразным привести цитату из руководства Э.Рупперта и соавторов "неопределенное систематическое положение турбеллярий (как и всего таксона Плоские черви) пока не удается прояснить, используя средства молекулярной систематики и компьютерной нумерической таксономии". Справедливости ради следует отметить, что этот же источник несколькими страницами раньше пишет "последние данные молекулярной систематики тоже свидетельствуют не в пользу традиционных взглядов" (под традиционными взгядами понимается морфологическая точка зрения, в частности планулоидная теория). Так что консенсус относительно происхождения турбеллярий остается не достигнутым, а студентам предлагается самостоятельно оценить доводы представленных гипотез.
Задание для самостоятельной работ: представьте себе, что свободно живущее животное начинает осваивать паразитический образ жизни. Определите, к каким новым факторам новой внешней среды этот организм должен приспособиться при переходе к паразитированию в кишечнике. Данное задание для самостоятельной работы является подготовкой студента к изучению адаптаций паразитов к их среде обитания. NB! При выполнении задания не следует забегать вперед и описывать адаптации паразитов.
ТЕМА 3.27
КЛАСС ТРЕМАТОДЫ ИЛИ СОСАЛЬЩИКИ (TREMATODA)
| Лиза повторила то, о чем она когда-то рассказывала Девешу: про ланцетовидного печеночного сосальщика, в чьем жизненном цикле участвуют три хозяина: травоядное животное, улитка и муравей. - И на определенном этапе сосальщик, по сути дела захватывает муравья, в котором обосновался. Он заставляет муравья взобраться по траинке, блокирует ему челюсти и ждет, когда его вместе с травой съест пасущаяся корова. Дж. Роллинс |
Как уже указывалось, огромное большинство представителей типа Плоские черви являются паразитами. Трематоды на разных фазах жизненного цикла паразитируют у разных хозяев: двуполое поколение трематод паразитирует у позвоночных, партеногенетические поколения - преимущественно приурочено к брюхоногим моллюскам. Типовой размер двуполого поколения трематод оценивается примерно в 10-12 миллиметров, есть виды меньших размеров - до 2 миллиметров, есть виды, достигающие в длину 5-7 сантиметров (печеночные двуустки). Остальные стадии жизненного цикла трематод характеризуются микроскопическими размерами. Общая численность трематод в настоящее время оценивается в руководстве Э.Рупперта с соавторами (2008) приблизительно в 11000 видов, а в руководстве Вестхайде и Ригера (2008) - в 7200 видов.
Трематоды как Плоские черви. Трематоды, а также другие паразитические представители плоских червей могут быть охарактеризованы как плоские черви, адаптированные к паразитизму. Главнейшие признаки типа Плоских червей у трематод выражены вполне отчетливо (лучше всего - у марит) и проявляются в общем плане строения тела: в форме тела, паренхиматозности, наличии замкнутой пищеварительной системы, ортогональной нервной системы, протонефридиальной выделительной системы, гермафодитизма (только одно семейство трематод - Кровяные двуустки - характеризуется раздельнополостью, причем этот признак явно является вторичным). Для всех паразитических плоских червей характерно также формирование сложного яйца (смотри раздел 3.20).
Общая схема жизненного цикла трематод. Трематоды от остальных плоских червей, в том числе и паразитических, отличаются сложным жизненным циклом, протекающим по типу гетерогонии. В жизненном цикле этих червей правильно сменяют друг друга гермафродитное поколение, размножающееся двуполым способом, приуроченное к позвоночным хозяевам - марита и два поколения, размножающихся партеногенетически и приуроченных к моллюскам - спороциста и редия (в жизненных циклах некоторых трематод место редии занимает дочерняя спороциста). В жизненном цикле трематод также присутствуют две личинки: мирацидий, который является личинкой первого партеногенетического поколения, и церкария, которая является личинкой гермафродитного поколения. В огромном большинстве жизненных циклов имеется также стадия инцистированной церкарии.
Гермафродитное (в случае кровяных двуусток - раздельнополое) поколение трематод обитает в позвоночных животных (окончательный хозяин). В результате двуполого размножения формируются "яйца", которые выходят во внешнюю среду. Выход "яиц" чаще всего совершается естественным путем, вместе с экскрементами, но если мариты локализованы не в кишечном тракте, то могут использоваться особые приемы, обеспечивающие выход "яиц". Иногда развитие зародыша начинается еще в матке материнского организма, так что к моменту выхода "яиц" из организма окончательного хозяина в них уже содержится сформированная личинка, но значительно чаще эти процессы происходят во внешней среде, но под оболочкой яйца, есть и промежуточные варианты, при которых развитие начинается в матке матери, а завершаентся во внешней среде. Как бы то ни было, в конечном итоге внутри яйца формируется стадия, которая называется мирацидий. Мирацидий выходит из яйца в воду. Мирацидий совершает активные поисковые движения и находит (если повезет) подходящего хозяина, в которого внедряется (активный путь заражения). Этого хозяина мы называем первым промежуточным хозяином. Таким хозяином практически у всех трематод являются моллюски, чаще всего брюхоногие. Мирацидий, внедрившись в хозяина, превращается в спороцисту. Превращение сопровождается морфологической перестройкой организации, таким образом, мирацидия мы должны оценить как личинку спорооцисты. Еще в организме мирацидия находилась группа рыхло расположенных клеток - зародышевых шаров, эти клетки в организме спороцисты начинают дробиться без оплодотворения. За дроблением следует дифференцировка клеток, и формируется следующая стадия - редия. Процесс формирования редий происходит без оплодотворения и поэтому оценивается как партеногенез, а спороциста может быть оценена как партеногенетическая самка. Редия также способная к партеногенезу, в результате которого формируется следующая стадия жизненного цикла - церкария. Церкарии покадают организм первого промежуточного хозяина. Церкарии активно плавают, а затем проникают в тело какого-либо вида животного и инцистируются. Эта последняя категория хозяев получает название второго промежуточного хозяина. Заражение окончательного хозяина осуществляется путем проглатывания инцистированнной лицинки. Для разных трематод характерен разный набор вторых промежуточных хозяев, но практически всегда эти хозяева являются типичной пищей окончательных хозяев. После проглатывания церкарий в организме окончательного хозяина превращается в мариту, то есть церкарий должен быть оценен как личинка мариты.
Для большей наглядности приведем общую схему жизненного цикла трематод.
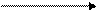
 марита
марита
двуполая стадия,
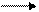 в окончательном хозяине=позвоночном
в окончательном хозяине=позвоночном
Церкария
личинка, во внешней среде,
позднее инцистируется
 "яйцо"
"яйцо"
во внешней среде
Редия мирацидий
партеногенетическая стадия, личинка, во внешней среде
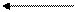 в первом промежуточном
в первом промежуточном
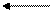 хозяине=моллюске
хозяине=моллюске
Спороциста
партеногенетическая стадия, в первом промежуточном хозяине=моллюске
Жизненные чиклы конкретных видов трематод могут отличаться от этой общей схемы. Главнейшими из вариантов являются следующие:
1. Яйцо из организма окончательного хозяина должно попасть не в воду, а в наземную среду, первым промежуточным хозяином являются наземные виды моллюсков, в этом случае вылупления мирацидиев не происходит, мирацидий как свободная стадий отсутствует (пример - ланцетовидная двуустка).
2. И у тех видов трематод, развитие которых связано с водой, в некоторых случаях мирацидий не вылупляется, заражение первого промежуточного хозяина происходит путем проглатывания яйца, то есть пассивным для трематоды путем (пример - кошачья двуустка, китайская двуустка).
3. В результате партеногенеза спороцисты формируется не редия, а стадия, которая называется дочерней спороцистой (пример - кровяная двуустка).
4. Церкарии могут не покидать первого промежуточного хозяина, заражение окончательного хозяина происходит при поедании первого промежуточного хозяина (пример Leucochloridium paradoxum). У этого вида трематод церкарии скапливаются в щупальцах янтарки (наземная улитка). Заражение окончательного хозяина - певчих птиц - может произойти при проглатывании моллюска целиком, или в том случае, если птица оторвет и проглотит только щупальце янтарки. Щупальца зараженного моллюска приобретают яркую окраску, что представляет пример демаскировки хозяина, выгодной для паразита.
5. Церкарии не подвергаются инцистированию, они отыскивают окончательного хозяина и внедряются в его теле перкутанно (пример - кровяная двуустка).
6. Церкарии могут инцистироваться на поверхности животных или растений, а не в организме второго промежуточного хозяина. Заражение окончательного хозяина проихсодит путем проглатывания инцистированного церкария вместе с пищевым объектом или за счет случайного проглатывания (пример - печеночная двуустка).
Характеристика стадий жизненного цикла трематод. Особенности организации марит трематод. Одной из особенностей трематод является наличие у них двух присосок, из которых первая помещается терминально на переднем конце тела, в глубине этой ротовой присоски находится рот. Вторая присоска называется брюшной, она размещена, понятно, на брюшной стороне тела, но положение ее у разных видов варьирует. Чаще всего она размещается на границе первой четверти или трети тела, но может быть размещена в задней половине тела и даже, например, у рода Diplodiscus (паразитирует в задней кишке лягушек) - терминально на заднем конце тела. Ранее полагали, что ротовое отверстие имеется и в брюшной присоске, с чем и связано название "двуустки", в учебно-научной литературе все чаще вместо термина "двуустки" пользуются термином "сосальщики".
Для трематод (а также и для всех остальных паразитических плоских червей) характерно наличие покрова особого типа. В русскоязычной литературе обычно покров такого типа называют тегументом, в аглоязычной - неодермисом. На определенной стадии развития у паразитических плоских червей присутствует типичный ресничный эпителий. В какой-то момент жизненного цикла этот покров сбрасывается и замещается выростами необластов (смотри раздел 3.20). При этом тела клеток, которые называются цитоны и в которых лежит ядро, остаются в паренхиме и непосредственно граничат с клетками паренхимы и мускульными клетками. Отростки необластов пронизывают базальную пластинку и разрастаются по ее внешней поверхности, причем отростки соседних необластов сливаются. В конечном итоге на поверхности тела формируется синцитиальный слой, лишенный ресничек. Этот слой может формировать шипики, которые полностью находятся внутри цитоплазмы, их основания прикрепляются к базальной пластинке, а вершины торчат наружу, выпячивая апикальную мембрану тегумента. Как и покровы других животных, тегумент (неодермис) выполняет защитную и опорную функции. В числе других функций тегумента следует назвать экскрецию, осморегуляцию и функцию поглощения веществ (за счет пиноцитоза). При интенсификации этой последней функции тегумент образует разнообразные выросты - микроворсинки. Специфическими для паразитов являются функции защиты от воздействия организма хозяина - иммунного воздействия и пищеварительных ферментов. Для мышечной системы характерно - в сравнении с турбелляриями - отсутствие диагональных мышц и в целом более слабое развитие.
Пищеварительная система начинается ротовым отверстием, которое расположено в ротовой присоске, Далее следует сосательная глотка, устроенная по типу бульбовидной глотки прямокишечных турбеллярий. За глоткой находится короткий пищевод, развевляюящийся на две слепо замкнутые ветви кишечника. В типовом случае эти две ветви доходят почти до заднего конца тела червя и более не ветвятся. У мелких видов ветви могут быть более короткими, доходя примерно до середины тела. Для крупных видов характерно, что две названные ветви в свою очередь сильно ветвятся. Это решает проблему транспортировки питательных веществ. Отметим, что у крупных видов ветвистую форму могут принимать гонады, а желточники представлены мелкими гроздевидными фолликулами практически у всех трематод. Это разветвеление органов увеличивает их поверхность, что опять-таки решает проблемы получения питательных веществ. Пищей трематодам служат разные субстанции тела хозяина: полупереваренное или полностью переваренное содержимое кишечника, клеточный детрит, тканевая жидкость, кровь. Переваривание пищи происходит в просвете кишечника. Все мариты трематод в той или иной степени поглощают растворенные питательные вещества поверхностью тегумента. У представителей семейства Strigeidae имеется особый бокаловидный орган, специализированный на внекишечном поглощении пищи. В редчайших случаях пищеварительная система отсутствует (род Anenterontrema).
Часть трематод полностью анаэробна, другие способны использовать кислород, в этом последнем случае часть энергетических потребностей организма удовлетворяется за счет аэробных, часть - за счет анаэробных процессов.
Нервная система устроена по общему ортогональному типу, число продольных стволов - 6, причем вентральные стволы развиты заметно сильнее латеральных и дорзальных. На переднем конце тела имеется нервный центр, который имеет вид широкого пояска, от него отходят вперед и назад упомянутые выше 6 стволов. Таким образом, нервный центр трематод представлен ортогональным мозгом. Сравнительно сильно развитые нервные сплетения, как и следовало ожидать, находятся в стенке глотки и в присосках. Органы чувств отсутствуют, но на поверхности тела расположены сравнительно немногочисленные сенсиллы.
Выделительная система протонефридиального типа. Канальцы протонефридиев сливаются в два главных канала, которые в свою очередь сливаются в непарный главный выделительный канал. В задней части этого канала находится расширение - мочевой пузырь. Формирование непарного выделительного ствола наблюдается у марит и их личинок церкарий, у мирацидиев, спороцист и редий слияния выделительных каналов не происходит.
Как уже указывалось, для трематод (кроме кровяных двуусток) характерен гермафродитизм Мужская половая система включает два семенника, два семяпровода, которые сливаются в общий проток. Этот проток, расширяясь, образует наружный семенной пузырек. Последний открывается в сумку цирруса (так называется особый тип совокупительного органа), в сумке находится циррус, внутренний семянной пузырек и предстательная железа. Открывается сумка цирруса в половую клоаку. В ходе полового акта циррус выворачивается.
Женская часть половой системы начинается яичником, от него отходит яйцевод. Рядом с яичником лежит семяприемник, открывающийся в яйцевод. По бокам тела мариты расположены гроздевидные желточники, их протоки также открываются в яйцевод. Яйцевод расширяется в особую камеру - оотип. Яйцеклетка выходит в яйцевод, туда же поступают из семяприемника сперма партнера. Оплодотворение происходит или в яйцеводе или в оотипе. В оотипе зигота получает сопровождение из желточных клеток, и этот комплекс одевается оболочкой, формируя сложное яйцо (в дальнейшем - "яйцо"). Таким образом, половая система трематод относится к неофорному типу, характерному и для части турбеллярий. Оболочка "яиц" формируется из веществ, выделяемых желточниками. От оотипа берет начало трубчатая матка, которая открывается в половую клоаку. В оотип открываются также отверстия многочисленных одноклеточных желез, которые в совокупности называются тельцем Мелиса. Жидкие выделения этих желез наполняют оотип и матку, создавая среду существования "яиц" (некоторые авторы указывают, что продукты выделения тельца Мелиса участвуют и в формировании скорлупы яиц). Имеется еще одна часть половой системы - Лауреров канал, он имеет вид короткого протока, отходящего от яйцевода или протока семяприемника. Лауреров канал открывается наружу на спинной стороне тела червя, из него выпускаются наружу излишки желточных клеток. Возможно, что Лауреров канал является редуцированным влагалищем.
Оплодотворение у гермафродитных видов перекрестное, но встречается и самооплодотворение. Циррус вводится в половую клоаку партнера и извергает сперму в отверстие матки. Сперма по матке проходит до оотипа, затем до протока семяприемника, в котором и запасается.
Вариации половой системы многочисленны, но незначительны и проявляются главным образом во взаиморасположении органов (семенники лежат позади яичника, семенники лежат впереди яичника и т.п).
Яйцо. Стадия яйца в особой характеристике не нуждается.
Мирацидий. Размеры мирацидиев составляют доли миллиметра (0.02 – 0.35). Тело мирацидия в поперечном сечении округлое. Передняя треть тела более широкая, задняя - округло сужается.Эта стадия покрыта ресничным эпителием, который сбрасывается при формировании спороцисты. Ресничный эпителий обеспечивает движения мирацидия. Пищеварительной системы нет, мирацидий потребляет запасы питательных веществ, вынесенные из яйца. Поэтому срок жизни мирацидия ограничен несколькими часами (9 – 24), но, во-первых, сроки различаются у разных видов, во-вторых, сроки жизни очень сильно зависят от условий среды: температура, количество кислорода, рН и других. Нервная система представлена рыхлым скоплением нервных клеток в передней части тела, от этого сплетения два ствола отходят вперед и два назад. У мирацидиев представлены органы чувств - глазки, устроенные по типу глазных бокалов и расположенные на переднем конце тела, и небольшие выросты – сосочки, которые являются органами хемо- и механорецепции. Для мирацидиев характерны направленные движения - таксисы. которые обеспечивают попадание мирацидиев в зону обитания хозяев. На переднем конце тела расположена группа желез, их секрет обеспечивает лизис покровов хозяина. На переднем же конце тела у многих видов расположены стилеты, обеспечивающие механическое разрушение покровов хозяина. Выделительная система представлена протонефридиями, имеется 2 или 4 терминальные клетки, два канальца и два выдедительных отверстия. В задней части тела находится группа недифференцированных клеток - зародышевых шаров.
Спороциста. Спороцисты обачно локализованы вблизи места внедрения мирацидия в хозяина, чаще всего в гемоцеле. Если мирацидий вылупляется из яйца в кишечнике хозяина, то спороцисты локализованы в стенке кишечника. Спороциста представляет собой тонкостенный мешок округловеретеновидной формы или более вытянутый, многие спороцисты формируют выросты, тогда их тело можно оценить как бесформенное. Размеры спороцист составляют 1-3 миллиметра, вытянутые спороцисты могут достигать в длину 6 -10 миллиметоров. Мышечные элементы развиты слабо или даже практически не выражены, животное практически неспособно к движению, хотя спороцисты некоторых видов совершают пульсирующие движения. Тегумент этой стадии несет много микроворсинок. Пищеварительная система полностью отсутствует. Спороцисты выделяют в среду своего обитания пищеварительные ферменты, расщепляющие органические вещества тканей хозяина, питание происходит за счет поглощения поверхностью тегумента растворенных веществ. Нервная система у спороцист не обнаруживается. Протонефридиальная система развита хорошо. В полости тела спороцисты (паренхимные элементы у спороцист отсутствуют) расположены зародышевые шары, из которых формируются особи следующего поколения. Совокупность зародышевых шаров называется диффузным яичником. Следующее поколение может быть представлено редиями или дочерними спороцистами. Потомство выходит из тела спороцисты через разрыв стенки ее тела. Спороцисты части видов имеют родильную пору, в этом случае потомство выходит через нее.
Дочерняя спороциста. По организации сходна с материнской прежде всего отсутствием кишечника, развитием системы микроворсинок и питанием через покровы, которые представлены тегументом с ворсинками. Размеры дочерних спороцист меньше, чем размеры материнских. Дочерние спороцисты имеют мускулатуру, подвижны, локализованы в печени моллюска. Ортогон представлен парой нервных стволов. Имеются немногочисленные сенсорные элементы. Протонефридии формируют два выделительных ствола, которые образуют общий непарный канал. Дочерние спороцисты, как и материнские, имеют диффузный яичник. В результате партеногенза формириуются церкарии. У некоторых видов поколение дочерних спороцист может произвести еще одно поколение спороцист, которые и сформируют церкарий. В стенке тела дочерних спороцист имеется отверстие - родильная пора, через которую потомство выходит наружу.
Редия. Для редий характерна вполне определенная форма тела, они червеобразны, в поперечном сечении округлы. В передней трети тела расположено кольцевое утолщение - воротничок, а задней - боковые выросты, так называемые латеральные крылья. Для небольшой части трематод характерны мешковидные и нитевидные редии без латеральных крыльев. Редии локализованы в печени или в полости тела хозяина. Тело редий одето тегументом с ворсинками, ворсинки развиты слабее, чем у спороцист. Мышечная система развита, редии подвижны. Имеется ротовое отверстие, слабо развитая глотка и мешковидный кишечник. Питание редий двойственно: через кишечник (при наблюдениями за редиями изредка отмечали даже каннибализм, более крупная редия может проглотить более мелкую) и через покровы, подобно спороцисте. Ортогон включает две пары нервных стволов, нервный центр имеет вид кольцевого пояска, расположенного непосредственно позади глотки. Протонефридиальная система развита хорошо. Терминальные клетки многочисленны, имеется два главных выделительных канала, которые сливаются в непарный канал. Редии, как и спороцисты, имеют диффузный яичник. В результате партеногенза формируются церкарии. У некоторых видов поколение редий может произвести еще одно поколение редий, которые и сформируют церкарий. В стенке тела редий имеется родильная пора, расположенная в районе воротничка. Продолжительность жизни редий составляет около 2.5 месяцев.
Церкария. Тело церкарий разделено на два отдела. Передний отдел - это собственно тело, задний - хвост. Тело церкарий имеет или плоскую листовидную или более или менее цилиндрическую форму. У церкарий большинства видов трематод длина хвоста приблизительно равна длине туловища. У части видов хвост значительно длиннее тела (в 7 – 8 раз). Размеры церкарий разных видов (общая длина тела с хвостом) составляют от 0.5 до 2 миллиметров. Хвост снабжен мощно развитой
мускулатурой, при сокращении мышц генерируются движение хвоста, обеспечивающее плавание церкарии.Хвост отбрасывается при инцистировании или проникновении церкарии в организм хозяина. Нервная система устроена принципиально так же, как и у марит. развита хорошо. У большинства видов имеются глазки. На поверхности тела расположены многочисленные хемо- и механорецепторы. Сочетание развитого рецепторного и двигательного аппарата обеспечивает поисковые движения церкарии. На переднем конце тела находится аппарат проникновения, представленный набором желез и - у части видов - стилетов. Церкарии имеет ротовую и брюшную присоски. Пищеварительная система развита, двуветвиста, но не функционирует. Церкарии, как и мирацидии, активно не питаются, в энергетических целях расходуется запасенный в их тканях гликоген. В связи с отсутствием активного питания церкарии характеризуются коротким сроком жизни (от 7-8 часов до 3-4 дней, сроки жизни различны у разных видов и, конечно, очень сильно зависят от условий среды). Протонефридиальная система развита хорошо, терминальные клетки многочисленны. Канальцы протонефридиев объединяются в непарный канал, который, однако, в их хвостике раздваивается, каждая из двух ветвей открывается наружу самостоятельным отверстием. Когда церкария теряет хвостик, у нее вторично образуется непарное выделительное отверстие. Церкарии являются личинками мариты. Они могут превращаться в мариту непосредственно. Этот вариант развития реализован у кровяных двуусток: церкарии, найдя подходящего хозяина, активно пробуравливают его покровы, попадают в кровеносную систему, где превращаются в марит. Чаще церкарии проникают в организм второго промежуточного хозяина, где инцистируется. Такая стадия называется метацеркарией. Метацеркария может попасть в кишечник окончательного хозяина при поедании им второго промежуточного хозяина. В кишечнике окончательного хозяина из метацеркарии формируется марита. Как уже указывалось, для некоторых трематод характерно, что церкарии инцистируются во внепшней среде, на повнерхности животных или растений. В этом случае инцистированную церкарию называют адолескарией. Для превращения в мариту адолескария должна быть проглочена соответствующим окончательным хозяином.
Уже указывалось, что процесс размножения спороцист и редий оценивается как партеногенез. Эта точка зрения не является всеобщей. Альтернативная точка зрения заключается в том, что процесс размножения спороцист и редий следует оценить как полиэмбрионию. Под полиэмбрионией понимается процесс, при котором зигота дает начало нескольким бластомерам, они теряют связь между собой, и каждый из них (или группа сцепленных бластомеров) развивается в новый организм. Однако для более примитивных трематод показано, что при формировании зародышевых шаров имеет место мейоз, что позволяет считать зародышевые шары яйцеклетками, а не зиготами и соответственно рассматривать процесс размножения трематод как партеногенез.
Завершая характеристику трематод, следует сказать, что эти животные по ходу жизненного цикла меняют среду обитания (внешняя среда, организм различных хозяев). Эти паразиты должны физиологически приспосабливаться к обитанию в разных условиях. Они по ходу жизненного цикла два или три раза должны найти подходящего хозяина и проникнуть в него. Приспособления трематод ко всем этим трудностям составляет их исключительно интересную биологию. Практическое значение плоских червей определяется тем, что они заражают экономически важных домашних и диких животных, а также человека. Например, кровяной двуусткой в настощее время заражено приблизительно 300 миллионов человек.
Жизненные циклы отдельных видов трематод изучаются студентами самостоятельно.
Наиболее распространенными трематодами человека являются печеночная двуустка - Fasciola hepatica, кошачья двуустка - Opisthorchis felineus, китайская двуустка Clonorchis sinensis, Paragonimus westermani (у этого вида тривиальное название отсутствует), кровяные двуустки - Schistosoma haematobium и S. mansoni. Редко встречается у человека, но часто у копытных животных ланцетовидная двуустка, для которой характерен удивительный цикл развития (особенности жизненного цикла ланцетовидной двуустки представлены в эпиграфе к настоящему разделу).
Печеночная двуустка известна с 18 века. Полный цикл ее развития был расшифрован в 1883 г., независимо друг от друга Лекартом и Томасом.
Кошачья, или сибирская двуустка была описана в 1891 г. К.Н.Виноградовым, промежуточный хозяин этой трематоды был обнаружен в 1932 г. Фогелем.
Заболевание, вызванное паразитированием Schistosoma, было известно еще в Древнем Египте. Представители вида Schistosoma haematobium были обнаружены в 1851 г. Бильгарцем. Позднее, в 1902 г. был описан вид Schistosoma mansoni (автор открытия - Manson).
Для конкретизации представлений студентов об особенностях морфологии и биологии трематод в таблице 3.27.1 приводятся некоторые дополнительные материала по нескольким видам трематод, встречающихся у человека. 1)
Табл. 3.27.1
Некоторые биологические и размерные характеристики
отдельных видов трематод
| Особенности строения и/или биологии | Fasciola hepatica 2) | Opisthorchis felineus 3) | Dicrocoelium lanceatum (dendriticum) | Schistosoma haematobium |
| Размеры мариты | длина 3 - 5 см, ширина - 8-18 мм | длина 8-13 мм ширина - 2-3 мм | длина 5 - 15 мм ширина 1.0 – 2.5 мм | самец: длина 18 - 20 мм, ширина - 1 мм. самка: длина - 10 - 14 мм, ширина - 0.25 мм |
| Продолжительность жизни мариты | от 10-12 месяцев до 3-5 лет. Известен случай продолжительности жизни 11 лет | свыше 3 лет, имеются сведения о продолжительности жизни 15 – 20- лет | несколько лет | 27 -28 лет, возможно и более |
| Продолжительность периода от заражения окончательного хозяина до начала выхода яиц | 3 - 4 месяца | 3 – 4 недели | 70 -80 дней | |
| Плодовитость мариты | около 1 млн. яиц в неделю | 160 – 190 яиц в сутки | ||
| Размеры "яйца" | 0.14 х 0.08 мм | 0.030 х 0.015 мм | 0.041 х 0.027 мм | 0.14 х -.0.05 мм. |
| Продолжительность эмбрионального развития | 2 -3 недели | |||
| Продолжительность жизни мирацидия в воде | 36 – 40 часов | Мирацидий вылупляется только после заглатывания яйца моллюском битинией – первым промежуточным хозяином. | Мирацидий вылупляется только после заглатывания яйца моллюском – первым промежуточным хозяином. | 24 часа |
| Продолжительность периода от заражения первого промежуточного хозяина до начала выхода церкариев | 30 – 70 дней | 4 – 6 месяцев | 3 – 12 недель | |
| Плодовитость партенитных стадий | общая численность потомства одной материнской спороцисты - 200 тысяч церкарий | |||
| Продолжительность периода миграции молодой двуустки к месту окончательной лакализации | 6 -7 недель |
Дата добавления: 2016-05-11; просмотров: 579;