КРАТКИЙ ОЧЕРК ИСТОРИИ АНАТОМИИ 52 страница
доли тянется под названием центральной части, pars centralis, которая на уровне заднего края мозолистого тела разделяется на нижний рог, cornu inferius, (в толще височной доли) и задний рог, cornu posterius (в затылочной доле).
Медиальная стенка переднего рога образована septum pellucidum, которая отделяет передний рог от такого же рога другого полушария (см. рис. 293). Латеральная стенка и отчасти дно переднего рога заняты возвышением серого цвета, головкой хвостатого ядра, caput nuclei caudati, а верхняя стенка образуется волокнами мозолистого тела. Крыша центральной, наиболее узкой части бокового желудочка также состоит из волокон мозолистого тела, дно же составляется из продолжения хвостатого ядра, corpus miclei caudati, и части верхней поверхности таламуса. Задний рог окружен слоем белых нервных волокон, происходящих из мозолистого тела, так называемого tapetum (покров); на его медиальной стенке заметен валик — птичья шпора, calcar avis, образованная вдавлением со стороны sulcus calcarinus, находящейся на медиальной поверхности полушария. Верхнелатеральная стенка нижнего рога образуется tapetum, составляющим продолжение такого же образования, окружающего задний рог. С медиальной стороны на верхней стенке проходит загибающаяся книзу и кпереди утонченная часть хвостатого ядра — cauda nuclei caudati.
По медиальной стенке нижнего рога на всем протяжении тянется белого цвета возвышение — гиппокамп, hippocampus, который образуется вследствие вдавления от глубоко врезывающейся снаружи sulcus hippocampi. Передний конец hippocampus разделяется бороздками на несколько небольших бугорков. По медиальному краю гиппокампа идет так называемая бахромка, fimbria hippocampi, представляющая продолжение ножки свода (crus fornicis). На дне нижнего рога находится валик, eminentia collateralis, происходящий от вдавления снаружи одноименной борозды. С медиальной стороны бокового желудочка в его центральную часть и нижний рог вдается мягкая мозговая оболочка, образующая в этом месте сосудистое сплетение, plexus choroideus ventriculi lateralis. Сплетение покрыто эпителием, представляющим остаток неразвитой медиальной стенки желудочка. Plexus choroideus ventriculi lateralis является латеральным краем tela choroidea ventriculi tertii.
Базальные ядра полушарий
Кроме серой коры на поверхности полушария, имеются еще скопления серого вещества в его толще, именуемые базальными ядрами и составляющие то, что для краткости называют подкоркой. В отличие от коры, имеющей строение экранных центров, подкорковые ядра имеют строение
Рис. 296. Фронтальный разрез полушарий через полосатое тело и таламус.
1 — fornix; 2 — plexus choroideus ventriculi tertii; 3 — plexus choroideus ventriculi lateralis; 4 — ventriculus lateralis; 5 — nucl. caudatus; 6 — crus posterius внутренней капсулы; 7 — putamen; 8 — globus pallidus; 9 — capsula externa; 10 — claustrum; 11 — sul. cerebri lateralis; 12, 14, 15 — gyri temporales superior, medius et inferior; 13 — insula; 16 — gyrus parahippocampalis; 17 — n. oculomotorius; 18 — pons; 19 — nucl. corporis mamillaris; 20 — tr. opticus; 21 — ventriculus tertius; 22 — adhesio interthalamica; 23, 24, 25 — ядра thalamus; 26, 28, 31 — gyri frontales inferior, medius et superior; 27 — corpus callosum; 29 — gyrus cinguii; 30 — sul. cinguii.
ядерных центров. Различают три скопления подкорковых ядер: corpus striatum, claustrum и corpus amygdaloideum (рис. 296, 297).
1. Coprus striatum, полосатое тело, состоит из двух не вполне отделенных друг от друга частей — nucleus caudatus и nucleus lentiformis.
A. Nucleus caudatus, хвостатое ядро, лежит выше и медиальнее nucleus lentiformis, отделяясь от последнего прослойкой белого вещества, называемой внутренней капсулой, capsula interna. Утолщенная передняя часть хвостатого ядра, его головка, caput nuclei caudati, образует латеральную стенку переднего рога бокового желудочка, задний же утонченный отдел хвостатого ядра, corpus et cauda nuclei caudati, тянется назад по дну центральной части бокового желудочка; cauda заворачивается на верхнюю стенку нижнего рога. С медиальной стороны niicleus caudatus прилегает к таламусу, отделяясь от него полоской белого вещества, stria terminalis. Спереди и снизу головка хвостатого ядра доходит до substantia perforata anterior, где она соединяется с niicleus lentiformis (с частью последнего, называемой putamen). Кроме этого широкого соединения обоих ядер с вентральной стороны, имеются еще тонкие полоски серого вещества, располагающиеся вперемешку с белыми пучками внутренней капсулы. Они послужили причиной названия «полосатое тело», corpus striatum (см. рис. 296).
Б. Nucleus lentiformis, чечевицеобразное ядро, залегает латерально от nucleus caudatus и таламуса, отделенное от них capsula interna. На горизонтальном разрезе полушария медиальная поверхность чечевицеобразного ядра, обращенная к внутренней капсуле, имеет форму угла с верхушкой, направленной к середине; передняя сторона угла параллельна хвостатому ядру, а задняя — таламусу. Латеральная поверхность немного
Рис. 297. Полушария большого мозга на разных уровнях горизонтального разреза.
 1 — nucl. caudatus; 2 — putamen; 3 — кора островка полушария; 4 — globus pallidus; 5 — claustrum; б — cauda nuclei caudati; 7 — nucl. corporis geniculati medialis; 8 — cornu inferius ventriculi lateralis; 9 — pedunculus cerebellaris superior; 10 — pedunculus cerebellaris medius; 11 — pedunculus cerebellaris inferior; 12 — striae medullares; 13 — trigonum n. hypoglossi; 14 — trigonum n. vagi;
1 — nucl. caudatus; 2 — putamen; 3 — кора островка полушария; 4 — globus pallidus; 5 — claustrum; б — cauda nuclei caudati; 7 — nucl. corporis geniculati medialis; 8 — cornu inferius ventriculi lateralis; 9 — pedunculus cerebellaris superior; 10 — pedunculus cerebellaris medius; 11 — pedunculus cerebellaris inferior; 12 — striae medullares; 13 — trigonum n. hypoglossi; 14 — trigonum n. vagi;
15 - tuberculum gracile; 16 — cerebellum; 17 — velum medullare superius; 18 — n. trochlearis; 19 — thalamus; 20 — nucl. ruber; 21 — stria terminalis; 22 — nucl. hypothalamicus; 23 — nucl. caudatus; 24 — cortex cerebri; 25 — cavum septi pellucidi; 26 — cornu anterius ventriculi lateralis; AA — capsula interna: a — fibrae corti- cothalamicae; 6 — tr. frontopontinus; в — tr. corticonuclearis; г — tr. corticospinalis; d — fibrae thalamocortica- les; e — tr. occipitotemporopontinus; ж — центральный слуховой тракт,
з — центральный зрительный тракт.
выпукла и обращена к латеральной стороне полушария в области островка. Спереди и вентрально, как было уже указано, чечевицеобразное ядро сливается с головкой nucleus caudatus. На фронтальном разрезе чечевицеобразное ядро имеет форму клина, верхушка которого обращена в медиальную сторону, а основание — в латеральную. Чечевицеобразное ядро двумя параллельными белыми прослойками, laminae medullares, разделяется на три членика, из которых латеральный, темно-серого цвета, называется скорлупой, putamen, а два медиальных, более светлых, носят вместе название бледного шара, globus pallidus (см. рис. 296).
Отличаясь уже по своему макроскопическому виду, globus pallidus имеет также и гистологическую структуру, отличную от других частей полосатого тела. Филогенетически globus pallidus представляет более старое образование (paleostriatum), чем putamen и nucleus caudatus (neostriatum).
Ввиду всех этих особенностей globus pallidus в настоящее время выделяют в особую морфологическую единицу под названием pallidum, тогда как обозначение striatum оставляют только за putamen и nucleus caudatus. Вследствие этого термин «чечевицеобразное ядро» теряет свое прежнее значение и может употребляться только в чисто топографическом смысле, а вместо прежнего названия corpus striatum хвостатое и чечевицеобразное ядро именуют стриопаллидарной системой. Стриопаллидарная система представляет собой главную часть экстрапирамидной системы (см. далее), а кроме того, она является высшим регулирующим центром вегетативных функций в отношении теплорегуляции и углеводного обмена, доминирующим над подобными же вегетативными центрами в hypothalamus.
2. Claustrum, ограда, представляет тонкую пластинку серого вещества, заложенную в области островка, между ним и putamen (см. рис. 297). От последнего она отделяется прослойкой белого вещества, capsula externa, а от коры островка — прослойкой, носящей название capsula extrema.
3. Corpus amygdaloideum, миндалевидное тело, расположено под putamen в переднем конце височной доли. Corpus amygdaloideum, по-видимому, относится к подкорковым обонятельным центрам и к лимбической системе. В нем оканчивается идущий из обонятельной доли и substantia perforata anterior пучок волокон, отмеченный при описании таламуса под названием stria terminalis (см. рис. 297).
Лимбическая система представляет комплекс образований конечного, промежуточного и среднего мозга, участвующий в регуляции различных вегетативных функций, поддержании постоянства внутренней среды организма (гомеостаза) и в формировании эмоционально окрашенных поведенческих реакций. Поэтому некоторые авторы обозначают лимбическую систему как «висцеральный мозг». Основную часть ее составляют структуры коры большого мозга, расположенные преимущественно на медиальной поверхности его полушарий, и тесно связанные с ними подкорковые образования, а именно: амигдалоидная область, конечная полоска, гипоталамус, гиппокамп, свод, септальная область, сосцевидные тела, сосцевидно-таламический пучок, таламус, поясная извилина. На медиальной поверхности полушарий большого мозга лимбическая система представлена поясной и парагиппокампальной извилинами.
Белое вещество полушарий
Все пространство между серым веществом мозговой коры и базальными ядрами занято белым веществом. Оно состоит из большого количества нервных волокон, идущих в различных направлениях и образующих проводящие пути конечного мозга. Нервные волокна могут быть разделены на три системы: 1) ассоциативные, 2) комиссуральные и 3) проекционные волокна.
А. Ассоциативные волокна (рис. 298) связывают между собой различные участки коры одного и того же полушария. Они разделяются на короткие и длинные. Короткие волокна, fibrae arcuatae cerebri, связывают между собой соседние извилины в форме дугообразных пучков. Длинные ассоциативные волокна соединяют более отдаленные друг^ от друга участки коры. Таких пучков волокон существует несколько. Cingulum, пояс,— пучок волокон, проходящий в gyrus fornicatus, соединяет различные участки коры gyrus cinguii как между собой, так и с соседними извилинами медиальной поверхности полушария. Лобная доля соединяется с нижней теменной долькой, затылочной долей и задней частью височной доли посредством fasciculus longitudinalis superior. Височная и затылочная доли связываются между собой через fasciculus longitudinalis inferior. Наконец, орбитальную поверхность лобной доли соединяет с височным полюсом так называемый крючковидный пучок, fasciculus uncinatus.
Б. Комиссуральные волокна, входящие в состав так называемых мозговых комиссур, или спаек, соединяют симметричные части обоих полушарий. Самая большая мозговая спайка — мозолистое тело, corpus callosum, связывает между собой части обоих полушарий, относящиеся к neencephalon.
Две мозговые спайки, commissiira anterior и commissura fornicis, гораздо меньшие по своим размерам, относятся к rhinencephalon и соединяют: commissura anterior — обонятельные доли и обе парагиппокампальные извилины, commissura fornicis — гиппокампы.












 В. Проекционные волокна связывают мозговую кору частью с thalamus и corpora geniculata, частью с нижележащими отделами центральной нервной системы до спинного мозга включительно. Одни из этих волокон проводят возбуждения центростремительно, по направлению к коре, а другие, наоборот,— центробежно.
В. Проекционные волокна связывают мозговую кору частью с thalamus и corpora geniculata, частью с нижележащими отделами центральной нервной системы до спинного мозга включительно. Одни из этих волокон проводят возбуждения центростремительно, по направлению к коре, а другие, наоборот,— центробежно.
Проекционные волокна в белом веществе полушария ближе к коре образуют так называемый лучистый венец, corona radiata, и затем главная часть их сходится во внутреннюю капсулу, о которой упоминалось выше. Внутренняя капсула, capsula interna, как было указано, представляет слой белого вещества между nucleus lentiformis, с одной стороны, и хвостатым ядром и таламусом — с другой. На фронтальном разрезе мозга внутренняя капсула имеет вид косо идущей белой полосы, продолжающейся в ножку мозга. На горизонтальном разрезе она представляется в форме угла, открытого в латеральную сторону (см. рис. 297); вследствие этого в capsula interna различают переднюю ножку, crus anterius capsulae internae, — между хвостатым ядром и передней половиной внутренней поверхности nucleus lentiformis, заднюю ножку, crus posterius, — между таламусом и задней половиной чечевицеобразного ядра и колено, genu capsulae internae, лежащее на месте перегиба между обеими частями внутренней капсулы. Проекционные волокна по их длине могут быть разделены на следующие системы, начиная с самых длинных:
1. Tractus corticospinalis (pyramidalis) проводит двигательные волевые импульсы к мышцам туловища и конечностей. Начавшись от пирамидных клеток коры средней и верхней частей предцентральной извилины и lobulus paracentralis, волокна пирамидного пути идут в составе лучистого венца, а затем проходят через внутреннюю капсулу, занимая передние две трети ее задней ножки, причем волокна для верхней конечности идут спереди волокон для нижней конечности. Далее они проходят через ножку мозга, pedunculus cerebri, а оттуда через мост в продолговатый мозг.
2. Tractus corticonuclearis — проводящие пути к двигательным ядрам черепных нервов. Начавшись от пирамидных клеток коры нижней части пред-
центральной извилины, они проходят через колено внутренней капсулы и через ножку мозга, затем вступают в мост и, переходя на другую сторону, оканчиваются в двигательных ядрах противоположной стороны, образуя перекрест. Небольшая часть волокон оканчивается без перекреста.
Так как все двигательные волокна собраны на небольшом пространстве во внутренней капсуле (колено и передние две трети задней ножки ее), то при повреждении их в этом месте наблюдается односторонний паралич (hemiplegia) противоположной стороны тела.
3. Tractus corticopontini — пути от мозговой коры к ядрам моста.' Они идут от коры лобной доли (tractus frontopontinus), затылочной (tractus occipitopontinus), височной (tractus temporopontinus) и теменной (tractus ра- rietopontinus). В качестве продолжения этих путей из ядер моста идут волокна в мозжечок в составе его средних ножек. При помощи этих путей кора большого мозга оказывает тормозящее и регулирующее влияние на деятельность мозжечка.
4. Fibrae thalamocorticalis et corticothalamici — волокна от таламуса к коре и обратно от коры к таламусу. Из волокон, идущих от таламуса, необходимо отметить так называемую центральную таламическую лучистость, которая является конечной частью чувствительного пути, направляющегося к центру кожного чувства в постцентральную извилину. Выходя из латеральных ядер таламуса, волокна этого пути проходят через заднюю ножку внутренней капсулы, позади пирамидного пути. Место это было названо чувствительным перекрестом, так как здесь проходят и другие чувствительные пути, а именно: зрительная лучистость, radiatio optica, идущая от corpus geniculatum laterale и piilvinar таламуса к зрительному центру в коре затылочной доли, затем слуховая лучистость, radiatio aciistica, направляющаяся от corpus geniculatum mediale и нижнего холмика крыши среднего мозга к верхней височной извилине, где заложен центр слуха. Зрительный и слуховой пути занимают самое заднее положение в задней ножке внутренней капсулы.
Морфологические основы динамической локализации функций
в коре полушарий большого мозга (центры мозговой коры)
Знание локализации функций в коре головного мозга имеет огромное теоретическое значение, так как дает представление о нервной регуляции всех процессов организма и приспособлении его к окружающей среде. Оно имеет и большое практическое значение для диагностики мест поражения в полушариях головного мозга.
Представление о локализации функций в коре головного мозга связано прежде всего с понятием о корковом центре. Еще в 1874 г. киевский анатом В. А. Бец выступил с утверждением, что каждый участок коры отличается по строению от других участков мозга. Этим было положено начало учению о разнокачественности коры головного мозга — цитоархитектонике (цитос — клетка, архитектонес — строю). В настоящее время удалось выявить более 50 различных участков коры — корковых цитоархитектони- ческих полей, каждое из которых отличается от других по строению и расположению нервных элементов. Из этих полей, обозначаемых номерами, составлена специальная карта мозговой коры человека (рис. 299).
По И. П. Павлову, центр — это мозговой конец так называемого анализатора. Анализатор — это нервный механизм, функция которого состоит в том, чтобы разлагать известную сложность внешнего и внутреннего мира на отдельные элементы, т. е. производить анализ. Вместе с тем благодаря широким связям с другими анализаторами здесь происходит и синтез,
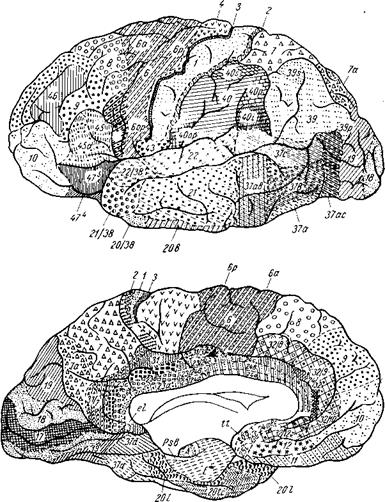 Рис. 299. Карта цитоархитектонических полей мозга человека (по данным Института мозга АМН СССР). Вверху — верхнелатеральная поверхность, внизу — медиальная поверхность. Объяснение в тексте.
Рис. 299. Карта цитоархитектонических полей мозга человека (по данным Института мозга АМН СССР). Вверху — верхнелатеральная поверхность, внизу — медиальная поверхность. Объяснение в тексте.
|
сочетание анализаторов друг с другом и с разными деятельностями организма. «Анализатор есть сложный нервный механизм, начинающийся наружным воспринимающим аппаратом и кончающийся в мозгу» (И. П. Павлов). С точки зрения И. П. Павлова, мозговой центр, или корковый конец анализатора, имеет не строго очерченные границы, а состоит из ядерной и рассеянной частей — теория ядра и рассеянных элементов. «Ядро» представляет подробную и точную проекцию в коре всех элементов периферического рецептора и является необходимым для осуществления высшего анализа и синтеза. «Рассеянные элементы» находятся по периферии ядра и могут быть разбросаны далеко от него; в них осуществляются более простой и элементарный анализ и синтез. При поражении ядерной части рассеянные элементы могут до известной степени компенсировать выпавшую функцию ядра, что имеет огромное клиническое значение для восстановления данной функции.
• До И. П. Павлова в коре различались двигательная зона, или двигательные центры, предцентральная извилина, и чувствительная зона, или чувствительные центры, расположенные позади sulcus centralis. И. П. Павлов показал, что так называемая двигательная зона, соответствующая пред- центральной извилине, есть, как и другие зоны мозговой коры, воспринимающая область (корковый конец двигательного анализатора). «Моторная область есть рецепторная область... Этим устанавливается единство всей коры полушарий» (И. П. Павлов).
В настоящее время вся мозговая кора рассматривается как сплошная воспринимающая поверхность. Кора — это совокупность корковых концов анализаторов. С этой точки зрения мы и рассмотрим топографию корковых отделов анализаторов, т. е. главнейшие воспринимающие участки коры полушарий большого мозга.
Прежде всего рассмотрим корковые концы анализаторов, воспринимающих раздражения из внутренней среды организма (см. рис. 289, 299).
1. Ядро двигательного анализатора, т. е. анализатора проприоцептивных (кинестетических) раздражений, исходящих от костей, суставов, скелетных мышц и их сухожилий, находится в предцентральной извилине (поля 4 и 6) и lobulus paracentralis. Здесь замыкаются двигательные условные рефлексы. Двигательные параличи, возникающие при поражении двигательной зоны, И. П. Павлов объясняет не повреждением двигательных эфферентных нейронов, а нарушением ядра двигательного анализатора, вследствие чего кора не воспринимает кинестетические раздражения и движения становятся невозможными. Клетки ядра двигательного анализатора заложены в средних слоях коры моторной зоны. В глубоких ее слоях (V, отчасти VI) лежат гигантские пирамидные клетки, представляющие собой эфферентные нейроны, которые И. П. Павлов рассматривает как вставочные нейроны, связывающие кору мозга с подкорковыми ядрами, ядрами черепных нервов и передними рогами спинного мозга, т. е. с двигательными нейронами. В предцентральной извилине тело человека, так же как и в задней, спроецировано вниз головой. При этом правая двигательная область связана с левой половиной тела и наоборот, ибо начинающиеся от нее пирамидные пути перекрещиваются частью в продолговатом, а частью в спинном мозге. Мышцы туловища, гортани, глотки находятся под влиянием обоих полушарий. Кроме предцентральной извилины, проприоцептивные импульсы (мышечно-суставная чувствительность) приходят и в кору постцентральной извилины.
2. Ядро двигательного анализатора, имеющего.отношение к сочетанному повороту головы и глаз в противоположную сторону, помещается в средней лобной извилине, в премоторной области (поле 5). Такой поворот происходит и при раздражении поля 17, расположенного в затылочной доле в соседстве с ядром зрительного анализатора. Так как при сокращении мышц глаза в кору мозга (двигательный анализатор, поле 8) всегда поступают не только импульсы от рецепторов этих мышц, но и импульсы от сетчатки (зрительный анализатор, поле 17), то различные зрительные раздражения всегда сочетаются с различным положением глаз, устанавливаемым сокращением мышц глазного яблока.
3. Ядро двигательного анализатора, посредством которого происходит синтез целенаправленных сложных профессиональных, трудовых и спортивных движений, помещается в левой (у правшей) нижней теменной дольке, в gyrus supramarginalis (глубокие слои поля 40). Эти координированные движения, образованные по принципу временных связей и выработанные практикой индивидуальной жизни, осуществляются через связь gyrus supramarginalis с предцентральной извилиной. При поражении поля 40 сохраняется способность к движению вообще, но появляется неспособность совершать целенаправленные движения, действовать — апраксия (праксия — действие, практика).
4. Ядро анализатора положения и движения головы — статический анализатор (вестибулярный аппарат) в коре мозга точно еще не локализован. Есть основания предполагать, что вестибулярный аппарат проецируется в той же области коры, что и улитка, т. е. в височной доле. Так, при поражении полей 21 и 20, лежащих в области средней и нижней височных извилин, наблюдается атаксия, т. е. расстройство равновесия, покачивание тела при стоянии. Этот анализатор, играющий решающую роль в прямохождении человека, имеет особенное значение для работы летчиков в условиях реактивной авиации, так как чувствительность вестибулярного аппарата на самолете значительно понижается.
5. Ядро анализатора импульсов, идущих от внутренностей и сосудов, находится в нижних отделах передней и задней центральных извилин. Центростремительные импульсы от внутренностей, сосудов, непроизвольной мускулатуры и желез кожи поступают в этот отдел коры, откуда исходят центробежные пути к подкорковым вегетативным центрам.
В премоторной области (поля 6 и 8) совершается объединение вегетативных и анимальных функций. Однако не следует считать, что только эта область коры влияет на деятельность внутренностей. На них оказывает влияние состояние всей коры полушарий большого мозга.
Нервные импульсы из внешней среды организма поступают в корковые концы анализаторов внешнего мира.
1. Ядро слухового анализатора лежит в средней части верхней височной извилины, на поверхности, обращенной к островку, — поля 41, 42, 52, где спроецирована улитка. Повреждение ведет к глухоте.
2. Ядро зрительного анализатора находится в затылочной доле — поля 17, 18, 19. На внутренней поверхности затылочной доли, по краям sulcus calcarinus, в поле 17 заканчивается зрительный путь. Здесь спроецирована сетчатка глаза, причем зрительный анализатор каждого полушария связан с полями зрения и соименными половинами сетчатки обоих глаз (например, левое полушарие связано с латеральной половиной левого глаза и медиальной правого). При поражении ядра зрительного анализатора наступает слепота. Выше поля 17 расположено поле 18, при поражении которого зрение сохраняется и только теряется зрительная память. Еще выше находится поле 19, при поражении которого утрачивается ориентация в непривычной обстановке.
3. Ядро обонятельного анализатора помещается в филогенетически древней части коры мозга, в пределах основания обонятельного мозга — uncus, отчасти гиппокампа (поле 11) (см. рис. 299, поля А и Е).
4. Ядро вкусового анализатора, по одним данным, находится в нижней части постцентральной извилины, близко к центрам мышц рта и языка, по другим — в uncus, в ближайшем соседстве с корковым концом обонятельного анализатора, чем объясняется тесная связь обонятельных и вкусовых ощущений. Установлено, что расстройство вкуса наступает при поражении поля 43.
Анализаторы обоняния, вкуса и слуха каждого полушария связаны с -рецепторами соответствующих органов обеих сторон тела.
5. Ядро кожного анализатора (осязательная, болевая и температурная чувствительность) находится в постцентральной извилине (поля 1,2,3) ив коре верхней теменной области (поля 5 и 7). При этом тело спроецировано в постцентральной извилине вверх ногами, так что в верхней ее части расположена проекция рецепторов нижних конечностей, а в нижней — проекция рецепторов головы. Так как у животных рецепторы общей чувствительности особенно развиты на головном конце тела, в области рта, играющего огромную роль при захватывании пищи, то и у человека сохранилось сильное развитие рецепторов рта. В связи с этим область последних занимает в коре постцентральной извилины непомерно большую зону. Вместе с тем у человека в связи с развитием руки как органа труда резко увеличились рецепторы осязания в коже кисти, которая стала и органом осязания. Соответственно этому участки коры, соответствующие рецепторам верхней конечности, много больше таковых нижней конечности. Поэтому, если в постцентральную извилину врисовать фигуру человека головой вниз (к основанию черепа) и стопами вверх (к верхнему краю полушария), то надо нарисовать громадное лицо с несообразно большим ртом, большую руку, особенно кисть с большим пальцем, резко превосходящим остальные, небольшое туловище и маленькую ножку. Каждая постцентральная извилина связана с противоположной частью тела вследствие перекреста чувствительных проводников в спинном и частью в продолговатом мозге.
Частный вид кожной чувствительности — узнавание предметов на ощупь — стереогнозия (стереос — пространственный, гнозис — знание) связана с участком коры верхней теменной дольки (поле 7) перекрестно: левое полушарие соответствует правой руке, правое — левой руке. При поражении поверхностных слоев поля 7 утрачивается способность узнавать предметы на ощупь, при закрытых глазах.
Описанные корковые концы анализаторов расположены в определенных областях мозговой коры, которая, таким образом, представляет собой «грандиозную мозаику, грандиозную сигнализационную доску» (И. П. Павлов). На эту «доску» благодаря анализаторам падают сигналы из внешней и внутренней среды организма. Эти сигналы, по И. П. Павлову, и составляют первую сигнальную систему действительности, проявляющуюся в форме конкретно-наглядного мышления (ощущения и комплексы ощущений — восприятия). Первая сигнальная система имеется и у животных. Но «в развивающемся животном мире на фазе человека произошла чрезвычайная прибавка к механизмам нервной деятельности. Для животного действительность сигнализируется почти исключительно раздражениями и следами их в полушариях большого мозга, непосредственно приходящими в специальные клетки зрительных, слуховых и других рецепторов организма. Это то, что и мы имеем в себе как впечатления, ощущения и представления от окружающей внешней среды, как общеприродной, так и от нашей социальной, исключая слово, слышимое и видимое. Это первая сигнальная система, общая у нас с животными. Но слово составило вторую, специально нашу сигнальную систему действительности, будучи сигналом первых сигналов...» (И. П. Павлов).
Таким образом, И. П. Павлов различает две корковые системы: первую и вторую сигнальные системы действительности, из которых сначала возникла первая сигнальная система (она имеется и у животных), а затем вторая — она имеется только у человека и является словесной системой. Вторая сигнальная система — это человеческое мышление, которое всегда словесно, ибо язык — это материальная оболочка мышления. Язык — это «...непосредственная действительность мысли» (Маркс К., Энгельс Ф. Соч., 2-е изд., т. 3, с. 448).
Путем весьма длительного повторения образовались временные связи между определенными сигналами (слышимые звуки и видимые знаки) и движениями губ, языка, мышц гортани, с одной стороны, и с реальными раздражителями или представлениями о них — с другой. Так, на базе первой сигнальной системы возникла вторая.
Дата добавления: 2016-06-24; просмотров: 592;