Строение мышечного волокна и механизм мышечного сокращения в норме и при патологии
Электрический стимул или нервный импульс, распространяясь по нейрону, достигает мышцы, которая в ответ на это раздражение отвечает, как правило, сокращением. Однако при патологии мышца, несмотря на раздражение, может не сокращаться, и наоборот, в отсутствии активации возможны ее сокращения. Эти обстоятельства повышают интерес к строению мышцы и механизму ее сокращения. Скелетные мышцы состоят из группы удлиненных многоядерных клеток, называемых мышечными волокнами. Мышечные волокна содержат миофибриллы (рис. 20).
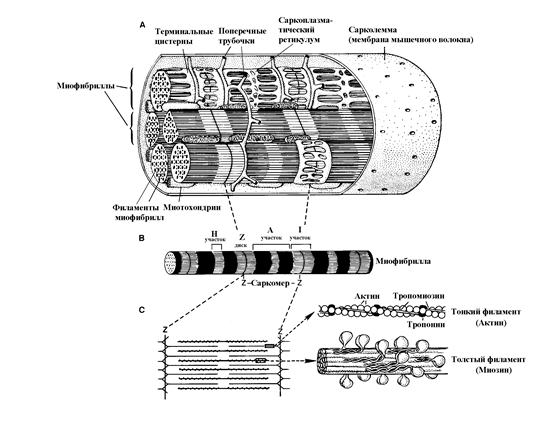
Рис. 20. Строение мышечного волокна и миофибрилл
(По W.Bloom and D.W.Faecett, 1975).
A – взаимоотношение миофибрилл, мембраны, тубулярной системы мышечного волокна,
B – саркомеры миофибриллы, разделенные Z-дисками,
C – взаимоотношения тонких и толстых филаментов в саркомере.
Мышечное волокно содержит до 1000 миофибрилл. Миофибрилла состоит из белковых нитей. Миофибриллы представлены повторяющимися по длиннику зонами - саркомерами. Саркомеры являются элементарной единицей мышечного сокращения и ограничены тонкой мембраной - Z зоной. Концы саркомера состоят из светлых участков I (изотропный - не обладает по своим оптическим свойствам двойным лучепреломлением), которые представлены более тонкими нитями белка актина. Центральная часть саркомера представлена участком А (анизотропный - по своим оптическим свойствам обладает двойным лучепреломлением), который состоит из светлой срединной части (Н) с более толстыми белковыми нитями (миозином) и периферической темной части с заходящими друг на друга белковыми нитями актина и миозина. Актиновые нити прикреплены к Z мембране, миозиновые нити расположены свободно, их концы накладываются на актиновые нити. Согласно теории скользящих филаментов Хаксли (H. Huxley, J. Hanson, 1954) при сокращении тонкие актиновые нити скользят вдоль миозиновых за счет того, что головки молекулы миозина прикрепляют себя к рецепторным местам молекул актина с образованием поперечных мостиков, способствующих смещению актиновых нитей вдоль миозиновых. При сокращении уменьшается в размерах I участок саркомера, а А участок остается неизменным. Миофибриллы завернуты в мешочную структуру, называемую саркоплазматическим ретикулумом, содержимое которого не сообщается ни с внутриклеточным пространством - цитоплазмой, содержащей миофибриллы, ни с внеклеточной жидкостью. Кроме саркоплазматического ретикулума в мышечном волокне имеется сеть поперечных трубочек (Т-трубочки), представляющих собой инвагинации мышечной мембраны и содержащие внеклеточную жидкость, которая не сообщается ни с саркоплазматическим ретикулумом, ни с цитоплазмой. Продольные трубочки саркоплазматического ретикулума вплотную соприкасаются с поперечными трубочками, образуя у поперечных трубочек терминальные цистерны. Активация мышечного сокращения инициируется возбуждением мембраны мышечного волокна. Возбуждение распространяется по поперечным трубочкам и переходит на терминальные цистерны, по которым оно распространяется вдоль мышечного волокна. Эта активация приводит к открытию потенциал–зависимых кальциевых каналов, через которые выходят ионы кальция, находящиеся в саркоплазматическом ретикулуме. Ионы кальция необходимы для активации рецепторов на молекулах актина для связывания их головками молекул миозина. После инициации сокращения кальций с помощью насоса переносится обратно в саркоплазматический ретикулум. Процесс актомиозинового взаимодействия, как и процесс активного обратного переноса ионов кальция в саркоплазматический ретикулум, происходит с использованием энергии, образуемой в результате расщепления АТФ. Таким образом, для возникновения мышечного сокращения необходима активация мышечной мембраны, эта активация возникает только при превышении критического уровня деполяризации в зоне концевой пластинки. Гиперполяризация мембраны, так же как и длительная деполяризация мембраны препятствует активации мембраны. Повышенная активация мышцы в условиях патологии может приводить к спазмам, которые обусловлены как неврогенными факторами, так и мышечными. Так болезненные спазмы, судороги при столбняке обусловлены снижением активности тормозных нейронов, повышенной активацией нейронов и частыми их разрядами, что приводит к сокращению мышечных волокон. Судороги при тетании обусловлены пониженным содержанием кальция в крови (экстрацеллюлярно), что способствует повышению проницаемости мембраны для ионов натрия, развитию деполяризации и частым разрядам нейронов, вызывающих спазм. При синдроме МакАрдля спазмы не связаны с уровнем активности нервной системы и обусловлены длительным нахождением кальция в цитоплазме мышечной клетки за счет дефицита АТФ, возникающего из-за фементативной (миофосфорилазной) недостаточности перевода гликогена в глюкозу. В клинических условиях коррекция повышенной мышечной возбудимости, мышечных спазмов (тетании) препаратами кальция (пероральное, парентеральное введение) направлена только на снижение уровня возбудимости нервных клеток. Коррекция кальция в сыворотке крови не способна влиять на кальциевые механизмы пресинаптического высвобождения ацетилхолина и кальциевые механизмы мышечного сокращения.
Механизм мышечного сокращения лежит в основе зависимости силы сократительной способности мышцы от ее исходной длины. Эндрью Хаксли показал, что чрезмерное укорочение, как и чрезмерное удлинение мышцы, снижает силу ее сокращения (рис. 21).
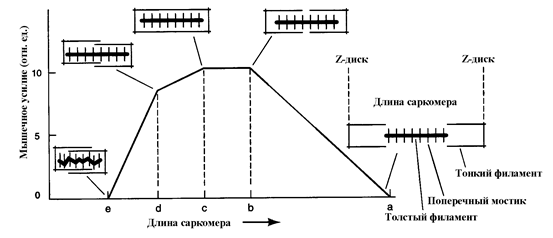
Рис. 21. Зависимость развиваемого усилия от исходной длины мышцы
(По Gordon et al., 1966. Адаптировано). Объяснение в тексте.
При чрезмерном укорочении мышцы схождение актиновых и миозиновых филаментов нарушает взаиморасположение головок молекул миозина и рецепторов актина (рис. 21, е), связи между ними формируются недостаточно прочные. Чрезмерное нефизиологическое растяжение мышцы приводит к расхождению актиновых и миозиновых миофиламентов (рис 21, а) и снижению усилия. В физиологических условиях возможно ослабление мышечного усилия при укорочении мышцы. Поэтому для создания максимального усилия, что важно в ЭНМГ диагностике больных с выраженными парезами, необходимо осуществлять произвольную активацию в умеренно растянутом состоянии мышцы.
Соответственно делению мотонейронов на малые и большие, в мышечной системе имеет место подразделение на мышечные волокна I типа (тонические) и мышечные волокна II типа (фазические). Это деление основано не только на скорости сокращения, но и на чувствительности к утомлению и особенностях обменных процессов (табл. 4). Волокна I типа используют аэробный окислительный путь энергопродукции (высокая активность окислительных ферментов), волокна II типа – гликолитические ферменты. Волокна второго типа подразделяются на 2 подгруппы: IIA – быстрые неутомляемые (FR) и IIB – быстрые утомляемые (FF).
Таблица 4
Сравнительная характеристика типов мышечных волокон
| Показатели | Тип I | Тип II A | Тип II B |
| Утомляемость | Низкая | Низкая | Высокая |
| Аэробный путь окисления | Высокий | Высокий | Низкий |
| Анаэробный путь окисления | Низкий | Высокий | Высокий |
| Скорость сокращения | Низкая | Высокая | Высокая |
| Сила сокращения | Низкая | Высокая | Высокая |
| Тип иннервирующих мотонейронов | Малые | Большие | Большие |
| Размер формируемых двигательных единиц | Малый | Большой | Большой |
| Диаметр иннервирующего аксона | Малый | Большой | Большой |
| Скорость проведения по иннервирующему аксону | Низкая | Высокая | Высокая |
| Порог рекрутирования иннервирующих мотонейронов | Низкий | Высокий | Высокий |
| Частота разрядов иннервирующих мотонейронов | Низкая | Высокая | Высокая |
| Частота миниатюрных потенциалов концевой пластинки | Низкая | Высокая | Высокая |
В связи с тем, что быстрые фазические мышечные волокна и соответствующие двигательные единицы имеют высокий порог активирования, они доступны исследованию методом игольчатой ЭМГ только при умеренном и выраженном напряжении мышц, так как включаются только вслед за медленными тоническими двигательными единицами, для активации которых достаточно легкое мышечное напряжение. Исследование СПИ по двигательным волокнам позволяет оценить проводимость по мотонейронам, формирующим быстрые двигательные единицы. Исследование же формы М-ответа позволяет учитывать состояние как быстрых, так и медленных двигательных единиц. Эти особенности, обусловленные различными типами мышечных волокон и двигательных единиц, необходимо учитывать при выборе методики исследования при различных видах патологии.
Дата добавления: 2016-03-27; просмотров: 3021;