Г л а в а 3. ДНК, РНК И СИНТЕЗ БЕЛКА
Простые органические молекулы, такие, как аминокислоты или нуклеотиды, ассоциируют с образованием больших полимеров. Две аминокислоты соединяются пептидной связью, два нуклеотида — фос-фодиэфирной. Последовательное повторение этих реакций ведет к образованию линейных полимеров, называемых соответственно полипептидами и полинуклеотидами. Полипептиды или белки и полинуклеоти-ды в форме ДНК и РНК считаются наиболее важными компонентами. Универсальные «кирпичики», из которых состоят белки, - это всего лишь 20 аминокислот, а молекулы ДНК и РНК построены только из четырех типов полинуклеотидов. Клетка содержит оба типа полинук-леотидов - ДНК и РНК; в ходе эволюции они специализировались и работают сообща, выполняя каждый свою функцию. Структура полинуклеотидов хорошо приспособлена для хранения и передачи информации. Химические различия между двумя типами полинуклеотидов делают их приспособленными для решения разных задач. Например, ДНК - хранилище генетической информации, так как ее молекула более стабильна, чем молекула РНК. Частично это обусловлено тем, что при наличии в РНК двух гидроксильных групп этот полинуклеотид в большей степени подвержен гидролизу.
Следовательно, вся информация о строении и функционировании любого живого организма содержится в закодированном виде в его генетическом материале, основу которого составляет ДНК. ДНК - длинная двухцепочечная полимерная молекула. В этой скрученной двойным жгутом гигантской молекуле «записаны» все признаки организма. Последовательность мономерных единиц (дезоксирибонуклеотидов) в одной ее цепи соответствует (комплементарна) последовательности дезоксирибонуклеотидов в другой. Принцип комплементарности обеспечивает идентичность исходных и новосинтезированных молекул ДНК, образующихся при удвоении (репликции).
Механизм комплементарного матричного копирования занимает центральное место в процессах переноса информации в биологических системах. Генетическая информация каждой клетки закодирована в последовательности оснований ее полинуклеотидов, и эта информация
передается из поколения в поколение благодаря комплементарное™ спаривания оснований.
Индивидуальными генетическими элементами со строго специфичной нуклеотидной последовательностью, кодирующими функциональные белки или РНК, являются гены. Гены находятся в ядре клетки, в хромосомах. В некоторых генах всего 800 пар нуклеотидов, в других -около миллиона. У человека 80-90 тыс. генов. Одни гены, называемые структурными, кодируют белки, другие - только молекулы РНК. Информация, содержащаяся в генах, которые кодируют белки, расшифровывается в ходе двух последовательных процессов: синтеза РНК, носящего название транскрипции и синтеза белка - трансляции. Сначала на определенном участке ДНК, как на матрице, синтезируется мРНК (информационная, матричная РНК) — в клетках животных этот процесс осуществляется в ядре. Затем, перенеся информацию из ядра в цитоплазму, в ходе согласованной работы многокомпонентной системы при участии тРНК (транспортных РНК), мРНК, ферментов и различных белковых факторов осуществляется синтез белковой молекулы. Все эти процессы обеспечивают правильный перевод зашифрованной в ДНК генетической информации с языка нуклеотидов на язык аминокислот. Аминокислотная последовательность белковой молекулы однозначно задает ее структуру и функции. Нуклеотиды как субъединицы ДНК, РНК выступают также в качестве переносчиков энергии.
Структура ДНК (рис. 5) - это линейный полимер. Его мономерные единицы (нуклеотиды) состоят из азотистого основания, пятиуглерод-ного сахара (пентозы) и фосфатной группы. Фосфатная группа присоединена к 5'-атому углерода моносахаридного остатка, органическое основание - к 1'-атому. Каждому нуклеотиду присвоено название, соответствующее названию входящего в его состав уникального основания. Основания в ДНК двух типов - пуриновые (аденин — А и гуанин — С) и пиримидиновые (цитозин - С, тимин - Т, урацил - U).
Нуклеотиды существовуют в двух оптических изомерах - L и D. Все без исключения живые организмы для построения своих нуклеотидов используют только D-формы. Присутствие даже малого количества L-формы нуклеотидов ингибирует или полностью блокирует работу ферментов синтеза ДНК.
В ДНК моносахарид представлен 2'-дезоксирибозой, содержащей одну гидроксильную группу, в РНК - рибозой, имеющей две гидро-ксильные группы. Нуклеотиды соединены друг с другом фосфодиэфир-ными связями, при этом фосфатная группа 5'- углеродного атома одного нуклеотида связана с 3’-ОН группой дезоксирибозы соседнего нуклеотида. На одном конце полинуклеотидной цепи находится 3’-ОН группа, на другом 5’-фосфатная группа.
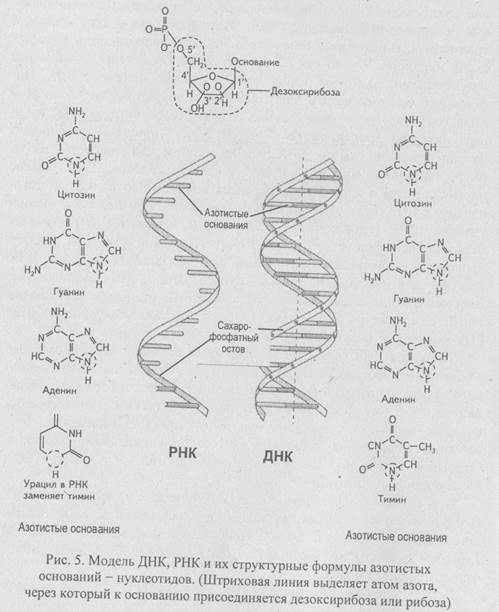
Нативная ДНК состоит из двух полимерных цепей, образующих спираль. Навитые одна на другую полинуклеотидные цепи удерживаются вместе водородными связями, образующимися между комплементарными основаниями противоположных цепей. При этом аденин образует пару только с тимином, гуанин - с цитозином. Пара оснований А-Т стабилизируется двумя водородными связями, пара С-С - тремя. Длина двухцепочечной ДНК обычно измеряется числом пар комплементарных нуклеотидов. Например, ДНК хромосомы 1 человека представляет собой одну двойную спираль длиной 263 миллиона пар нуклеотидов.
Сахарофосфатный состав молекулы, состоящий из фосфатных групп и дезоксирибозных остатков, соединенных 5'—З'-фосфодиэфирными связями, образует «боковины винтовой лестницы», а пары А-Т и С-С - «ее ступеньки». Цепи молекулы ДНК антипаралельны: одна из них имеет направление 3'—5', другая 5'—>3'. Нуклеотиды считают парами потому, что в молекуле ДНК две цепочки и их нуклеотиды соединены попарно поперечными связями.
Носитель генетической информации должен удовлетворять двум требованиям — воспроизводиться (реплицироваться) с высокой точностью и детерминировать (кодировать) синтез белковых молекул. Согласно принципу комплементарности, каждая цепь ДНК может служить матрицей для образования новой комплементарной цепи. Когда клетке необходимо разделиться, непосредственно перед этим она копирует молекулу ДНК в своих рибосомах. При этом две нити ДНК расходятся и на каждой из них, как на матрице, собирается дочерняя нить, в точности повторяющая ту, что была соединена с данной нитью в родительской клетке. В итоге появляются две идентичные дочерние хромосомы, которые при делении распределяются по разным клеткам. Так происходит передача наследственных признаков от родителей потомкам у всех клеточных организмов, имеющих ядро. Следовательно, после каждого раунда репликации образуются две дочерние молекулы, каждая из которых имеет такую же нуклеотидную последовательность, как исходная молекула ДНК. Нуклеотидная последовательность структурного гена однозначно задает аминокислотную последовательность кодируемого ею белка. Следовательно, каждая цепь ДНК служит матрицей при синтезе новой комплементарной цепи, а последовательность оснований в синтезируемой (растущей) цепи задается последовательностью комплементарных оснований цепи-матрицы.
Синтез ДНК у про- и эукариот осуществляется при участии множества различных ферментов. Основную роль играет ДНК-полимераза, которая последовательно присоединяет звенья растущей полинуклеотидной цепи в соответствии с принципом комплементарности и катализирует образование фосфодиэфирных связей.
Для разделения ДНК разработаны специальные гели на основе агарозы (полисахарид, выделяемый из морских водорослей). Предложена модификация гельэлектрофореза в агарозном геле, названная пульс-электрофорез, позволяющая разделять большие молекулы ДНК.
Определены последовательности нуклеотидов генов многих млекопитающих, включая гены, кодирующие гемоглобин, инсулин, цитохром С. Объём информации о ДНК столь велик (многие миллионы нуклеотидов), что для хранения и анализа имеющихся данных необходимы мощные компьютеры.
Для определения того, какие гены активны в данном типе клеток (идентификация специфических последовательностей), используют метод, именуемый ДНК-футпринтинг. Фрагменты ДНК метят Р , затем расщепляют нуклеазами, разделяют на геле и выявляют на радиоавтографе. Если водный раствор ДНК нагреть до 100 °С и сильно защело-чить (рН 13), то комплементарные пары оснований, удерживающие две цепи двойной спирали вместе, разрушаются и ДНК быстро диссоциирует на две цепи. Этот процесс, называемый денатурацией ДНК, ранее считался необратимым. Но если комплементарные цепи ДНК выдержать при температуре 65 °С, они легко спариваются, восстанавливая структуру двойной спирали, - процесс получил название ренатурации.
Подавляющее большинство генов содержит в закодированном виде информацию о синтезе белков. Полипептидам присуща большая универсальность, они состоят из аминокислот с химически разнообразными боковыми цепочками и способны принимать разные пространственные формы, которые насыщены реакционноспособными участками. Свойства полипептидов делают их идеально подходящими для выполнения разнообразных структурных и функциональных задач. Белки участвуют практически во всех процессах, протекающих в живых системах, они служат катализаторами биохимических реакций, осуществляют транспорт внутри и между клетками, регулируют проницаемость клеточных мембран, из них строятся различные структурные элементы. Белки - не только основной строительный материал живого организма, многие из них - ферменты, управляющие процессами в клетке. Белки участвуют в осуществлении двигательных функций, обеспечивают защиту от инфекций и токсинов, регулируют синтез остальных генных продуктов.
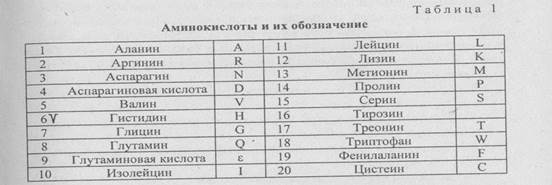
Все аминокислоты имеют сходное химическое строение: к центральному атому углерода присоединен атом водорода, аминогруппа, карбоксильная группа и боковая цепь. Существует 20 разных боковых групп и соответственно 20 аминокислот: например, в аминокислоте аланин боковой цепью является метильная группа (табл. 1).
Пептидная связь образуется между карбоксильной группой одной аминокислоты и аминогруппой другой. Первая аминокислота белковой молекулы имеет свободную аминогруппу (N-конец), последняя - свободную карбоксильную группу (С-конец).
Длина белковых молекул варьирует от 40 до 1000 аминокислотных остатков; в зависимости от их последовательности и аминокислотного состава молекулы белков принимают разную форму (конфигурацию, конформацию). Многие функционально активные белки состоят из двух и более полипептидных цепей, как идентичных, так и несколько различающихся. Белки, выполняющие ключевые функции, представляют собой сложные белковые комплексы, состоящие из множества разных полипептидных цепей - субъединиц.
С помощью генетического кода полинуклеотидная последовательность определяет последовательность аминокислот в белке; различные триплеты нуклеотидов кодируют специфические аминокислоты.
Важное «передаточное звено» при переводе генетической информации с языка нуклеотидов на язык аминокислот - РНК (рибонуклеиновые кислоты), которые синтезируются на определенных участках ДНК, как на матрицах, в соответствии с их нуклеотидной последовательностью.
Молекулы РНК несут информацию, они обладают химической индивидуальностью, влияющей на их поведение. Молекула РНК обладает двумя важными свойствами: закодированная в её нуклеотидной последовательности информация передаётся в процессе репликации, а уникальная пространственная структура определяет характер взаимодействия с другими молекулами и реакцию на внешние условия. Оба этих свойства - информационное и функциональное - являются необходимыми предпосылками эволюционного процесса. Нуклеотидная последовательность молекулы РНК аналогична наследственной информации, или генотипу организма. Пространственная укладка аналогична фенотипу - совокупности признаков организма, подверженного действию естественного отбора.
РНК (рис. 5) — линейная полинуклеотидная молекула, отличающаяся от ДНК по двум параметрам:
1. Моносахаридом в РНК является рибоза, содержащая не одну а две гидроксильные группы;
2. Одним из четырех оснований в РНК является урацил, занимающий место тимина.
Существование РНК в виде одной нити обусловлено:
отсутствием у всех клеточных организмов фермента для катализа реакции образования РНК на матрице РНК; такой фермент есть лишь у некоторых вирусов, гены которых «записаны» в виде двухнитчатой РНК, остальные организмы могут синтезировать молекулы РНК только на ДНК-матрице; из-за отсутствия метильной группы у урацила связь между аде-нином и урацилом малоустойчива и «удержание» второй (комплементарной) нити для РНК является проблемным. По причине однонитчатости РНК, в отличие от ДНК, не закручивается в спираль, а образует структуры в виде «шпилек», «петель». Спаривание оснований в молекуле РНК происходит таким же образом, как и в ДНК, за исключением того, что вместо пары А-Т, образуется А-U Комплементарные основания, как и в ДНК, соединены между собой водородными связями.
Существуют три основных типа РНК:
информационная (мРНК);
рибосомная (рРНК);
транспортная (тРНК).
Правильность транскрипции, т.е. ее начало и завершение в нужных сайтах (специфических участках), обеспечивают специфические нук- -леотидные последовательности в ДНК, а также белковые факторы. Транскрипция на ДНК осуществляется в клеточном ядре. Молекулы мРНК переносят информацию из ядра в цитоплазму, где она используется при трансляции белков, аминокислотные последовательности которых закодированы в последовательностях нуклеотидов мРНК (т.е., в конечном счете, в ДНК). мРНК связана с рибосомами, в которых осуществляется соединение аминокислот с образованием белков. Рибосомы - нуклеотидные частицы, в состав которых входит высокополимерная РНК и структурный белок. Биохимическая роль рибосом - синтез белка. Именно на рибосомах происходит соединение отдельных аминокислот в полипептиды, завершающееся образованием белков.
У большинства прокариот транскрипция всех РНК осуществляется с участием одной и той же РНК-полимеразы. У эукариот мРНК, рРНК, тРНК транскрибируются разными РНК-полимеразами.
С генетической точки зрения ген представляет собой специфическую нуклеотидную последовательность, траскрибируемую в РНК. Большинство транскрибируемых последовательностей ДНК составляют структурные гены, на которых синтезируется мРНК. Конечным продуктом структурного гена является белок. У прокариот структурный ген представляет собой непрерывный участок молекулы ДНК. У эукариот большинство структурных генов состоит из нескольких дискретных (отдельных) кодирующих областей — экзонов,разделенных некоди-рующими областями - нитронами.По завершении транскрипции эука-риотического структурного гена интроны вырезаются ферментами из первичного продукта транскрипции, экзоны сшиваются друг с другом «торец в торец» (сплайсинг)с образованием мРНК. Обычно длина экзонов составляет от 150 до 200 нуклеотидов, длина интронов варьирует от 40 до 10000 нуклеотидов.
В активно функционирующей клетке примерно 3-5% суммарной РНК приходится на долю мРНК, 90% — на долю рРНК, 4% — на долю тРНК. мРНК может быть представлена десятками различных типов молекул; рРНК - двумя типами. Более крупная рРНК образует с белками рибонуклеотидный комплекс, называемый большой рибосомной субъединицей. рРНК меньшего размера — комплекс, называемый малой ри-босомальной субъединицей. При синтезе белков субъединицы объединяются с образованием рибосомы. рРНК принадлежит роль главного катализатора в процессе синтеза белка, она составляет более 60% массы рибосомы. В эволюционном аспекте рРНК представляет собой основной компонент рибосомы.
Помимо тысяч рибосом в клетке, активно синтезирующей белки, содержится до 60 различных видов тРНК. тРНК - это линейная одноце-почечная молекула длиной от 75 до 93 нуклеотидов, имеющая несколько взаимно комплементарных участков, спаривающихся между собой. С помощью специфических ферментов (аминоацил-тРНК-синтетаз) к 3'-концу тРНК присоединяется соответствующая аминокислота. Для каждой из 20-ти аминокислот, из которых состоят все белки, существует, по крайней мере, одна специфическая тРНК. На другом конце молекул тРНК расположена последовательность из трех нуклеотидов, называемая антикодоном, она распознает специфический кадок в мРНК и определяет, какая аминокислота будет присоединена к растущей полипептидной цепи.
Трансляция (синтез белка) осуществляется при участии мРНК, разных тРНК, «нагруженных» соответствующими аминокислотами, рибосом и множества белковых факторов, обеспечивающих инициацию, элонгацию, терминацию синтеза полипептидной цепи.
Нуклеотидная последовательность, в которой закодировано более одного белка, называется опероном. Оперон находится под контролем единственного промотора, и при его транскрипции образуется одна длинная молекула мРНК, кодирующая несколько белков.
Синтез мРНК и соответственно синтез белка строго регулируется, так как у клетки недостаточно ресурсов для одновременной транскрипции и трансляции всех структурных генов. Про- и эукариоты постоянно синтезируют только те мРНК, которые необходимы для выполнения основных клеточных функций. Экспрессия остальных структурных генов осуществляется под строгим контролем регуляторных систем, запускающих транскрипцию только в случае возникновения потребности в определенных белках. За включение и выключение транскрипции отвечают дополнительные факторы транскрипции, которые связываются с соответствующими участками ДНК.
При синтезе белковых молекул первичной стадией образования полипептидной цепи белка является процесс активации аминокислот с помощью аденозинтрифосфата. Процесс активации идет при участии ферментов, в результате чего образуются аминоациладенилаты. Затем под действием фермента аминоацил-тРНК-синтетазы (для каждой из 20 аминокислот имеется свой особый фермент) «активированная» аминокислота соединяется с тРНК. Далее комплекс аминоацил-тРНК переносится на рибосомы, где происходит синтез полипептида. Пептидная связь образуется между карбоксильной группой одной аминокислоты и аминогруппой другой. Первая аминокислота белковой молекулы имеет свободную аминогруппу (N-конец), последняя - свободную карбоксильную группу (С-конец).
Сформировавшиеся белки освобождаются из рибосом, а рибосомы после этого могут присоединять новые комплексы аминоацил-тРНК и синтезировать новые белковые молекулы. Рибосомы связаны с мРНК, которая определяет последовательность чередования аминокислот в полипептидных цепочках. Таким образом, целостность и функциональная активность рибосом в клетках - одно из необходимых условий синтеза белковых молекул.
Тест-контроль к главе 3 Выберите правильные ответы:
1. Утверждение «ДНК является хранилищем генетической информации, потому, что ее молекулы в отличие от РНК более стабильны»:
А - верно;
Б - не верно;
В - требует уточнения.
2. Носитель генетической информации должен удовлетворять требованиям:
А — реплицироваться с высокой точностью;
Б - не подвергаться химическому гидролизу;
В - детерминировать синтез белковых молекул;
Г - выступать в качестве переносчика энергии;
Д - образовывать замкнутую кольцеобразную структуру.
3. Для разделения молекул ДНК используют:
А — высаливание;
Б - обратный осмос;
В - пульс-электрофорез;
Г - гельэлектрофорез;
Д - электродиализ.
4. Отличие молеклы РНК от молекулы ДНК:
А - моносахаридом является дезоксирибоза;
Б - моносахаридом является рибоза;
В - азотистое основание - тимин;
Г — азотистое основание — урацил;
Д — азотистое основание — гуанин.
5. Синтез молекулы ДНК осуществляется:
А - ДНК-лигазой;
Б - ДНК-полимеразой;
В — из L-формы нуклеотидов;
Г - из D-формы нуклеотидов;
Д - из смеси Dи L-форм нуклеотидов.
6. Сплайсинг:
А — вырезание из предшественника мРНК экзонов и ковалентное соединение интронов с образованием зрелых молекул мРНК;
Б - вырезание из предшественника мРНК интронов и ковалентное соединение экзонов с образованием зрелых молекул мРНК;
В - синтез зрелых молекул тРНК из путем сшивки отдельных нук-леотидов «торец в торец»;
Г — вырезание из предшественника мРНК интронов и их ковалентное соединение с образованием зрелых молекул мРНК;
Д - последовательное ковалентное соединение экзонов и интронов с образованием зрелых молекул мРНК.
7. Ко дон:
А -три соседних нуклеотида мРНК, кодирующих определенную аминокислоту;
Б - три соседних нуклеотида тРНК, комплементарный нуклеотидам специфического кодона в молекуле мРНК;
В -три соседних нуклеотида тРНК, кодирующих определенную аминокислоту;
Г — три соседних нуклеотида тРНК, кодирующих определенную последовательность аминокислот;
Д -три соседних нуклеотида мРНК, кодирующих определенную аминокислоту.
8. Уникальная пространственная структура молекулы РНК определяет:
А - процесс репликации;
Б — генотип;
В — фенотип;
Г - характер взаимодействия с другими молекулами и внешними
условиями; Д - локализацию молекулы РНК.
9. Процессы транскрипции идут:
А - постоянно с одинаковой скоростью;
Б — под контролем регуляторных систем;
В - периодически по мере накопления энергии;
Г — сопряжено с процессами формирования молекул ДНК;
Д -со скоростью, пропорциональной формированию структурных генов.
10. Оперон:
А - участок ДНК, содержащий несколько структурных генов;
Б - участок ДНК, содержащий один структурный ген;
В - нуклеотидная последовательность, кодирующая один белок;
Г - нуклеотидная последовательность, кодирующая более одного
белка;
Д - длинная молекула мРНК, кодирующая несколько белков.
Дата добавления: 2016-02-24; просмотров: 4556;