ОРГАНЫ ДЫХАНИЯ, ГАЗООБМЕН
У рыб в процессе эволюции развились два типа дыхания: водное И воздушное Водное дыхание осуществляется при помощи жабр и ; кожи, воздушное дыхание — при помощи кожи, плавательного пу-г даря, кишечника и наджаберных органов. Основными органами дакания рыб являются жабры, а все остальные можно назвать до-ршштельными или вспомогательными органами дыхания, хотя » некоторые из них иногда имеют первостепенное значение. ! Главной функцией жабр является газообмен — поглощение кис-•> $орода и выделение углекислого газа (диоксида углерода), но жабры участвуют также в водно-солевом обмене, выделяя аммиак, мочеви-11|у, поглощая и выделяя воду и ионы солей, особенно ионы натрия. | \" Устройство жаберного аппарата у рыб разных систематических И>упп существенно различается У круглоротых жабры мешковид-'е Йые, у хрящевых — пластинчатые, у костистых — гребенчатые. » Жабры (branchiae) костистых рыб расположены в боковых отде-, Дах жаберной полости, прикрытой с обеих сторон жаберными . крышками. Они состоят из четырех полных дут и пятой рудимен-тарованной жаберной дуги. От внешней выпуклой стороны дуги бтходят жаберные лепестки, основу которых составляют хрящевые жаберные лучи. Своим основанием лепестки прикрепляются к жа-Верной дуге, а свободные концы их расходятся под острым углом Наружу и внутрь (рис. 12) Поперек жаберного лепестка расположены тонкие респираторные складки, называемые вторичными пластинками, или лепесточками. Число лепесточков у разных видов рыб колеблется от 15 до 36 на 1 мм, их высота 150-200 мкм, толщина 10—15 мкм.
На стороне дуги, обращенной в ротовую полость, находятся жаберные тычинки. Они являются фильтрующим аппаратом, задерживающим во рту кормовые организмы и взвешенные частицы. У планктонофагов тычинки густые, высокие; у хищников — редкие, низкие; у всеядных занимают среднее положение между первыми и вторыми. Основу тычинок составляют костные пластинки.

|
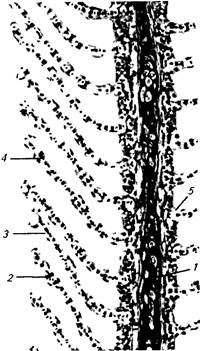
Рис 12 Жаберный аппарат костистой
рыбы (по Воскобойникову) 1 — первая жаберная дуга, 2 — жаберные тычинки, 3 — сердце, 4 — жаберные лепестки
В жабрах происходит интенсивное кровообращение, поэтому для них характерна ярко-розовая окраска Венозная кровь поступает в жабры по брюшной аорте, от которой к каждой жаберной дуге отходят бранхиальные артерии. От последних ответвляются приносящие лепестковые артерии, идущие по внутреннему краю хрящевых лучей От них к лепесточкам отделяются веточки, которые, раздваиваясь, идут по их наружному и внутреннему краям и образуют густую сеть капилляров. Диаметр капилляров приблизительно равен величине эритроцита. На противоположном конце лепесточка краевые артериолы сливаются в один сосуд, который впадает в выносящую вену, а последняя переходит в бранхиальные вены, впадающие в спинную аорту.
Гистологическое строение лучше изучать на продольных срезах, проходящих по ходу жаберных лепестков и параллельно жаберным дугам. В этом случае лепестки и тычинки будут разрезаны вдоль, а на поперечном сечении видны респираторные складки (рис. 13). Поверхность тычинок, дуг и лепестков в участках, свободных от лепесточков, покрыта многослойным эпителием эпидермального типа, содержащим многочисленные слизистые клетки. Между складками у их основания, кроме того, располагаются светлые хло-ридные клетки, участвующие в осморегуляции.
Респираторные складки снаружи выстланы однослойным плоским (респираторным) эпителием, а внутри пронизаны многочисленными капиллярами, которые поддерживаются опорными клетками (пилар-клетками). Следовательно, тканевый барьер между водой и кровью достаточно тонкий, образованный дыхательным эпителием и стенкой капилляров. При воздействии вредных факторов среды, особенно токсических веществ, он сильно утолщается за счет отслоения, набухания и пролиферации эпителия, что затрудняет диффузию кислорода в кровь и приводит к гипоксии.
Газообмен между водой и кровью рыб происходит путем пассивной диффузии кислорода и диоксида углерода по градиенту концентрации газов, только движение их идет в противоположных направлениях. Важнейшими условиями оптимального газообмена являются постоянная проточность воды в жабрах, сохранение их структуры, регулирование площади и плотности диффузионной
поверхности и использование гемоглобина для увеличения кислородной емкости плазмы крови. У рыб кровь в капиллярах жабр течет в направлении, противоположном току воды. Этот противоточный щеханизм обеспечивает практически полное насыщение крови кислородом и максимальное его извлечение из воды (у многих рыб до 80 % растворенного в воде кислорода).
Вода прокачивается через жаберную полость с помощью движения ротового аппарата и жаберных крышек. Поэтому частоту дыхания у рыб определяют по количеству движений жаберных крышек. На ритм дыхания рыб в первую очередь влияет содержание в воде кислорода, а также концентрация диоксида углерода, температура, рН и т. д. Причем чувствительность рыб к недостатку кислорода (в воде и крови) значительно выше, чем к избытку диоксида углерода (гиперкапния). Например, при 10 °С и нормальном содержании кислорода (4,0—5,0 мг/л) форель совершает 60—70, карп — 30—40 дыхательных движений в минуту, а при 1,2 мг О2/л частота дыхания увеличивается в 2—3 раза. Зимой у карпа ритм дыхания резко замедляется (до 3—4 дыхательных движений в минуту).
|
Большинство рыб, периодически прикрывая жаберные крышки, обратным током воды промывают жабры от взвесей. Это называют рефлексом откашливания или кашлем рыб. Он особенно четко проявляется при различных токсикозах.
Жабры помимо дыхательной функции выполняют функции Выделения и осморегуляции. Рыбы как водные животные относятся к аммониотелическим Организмам, у которых основным конечным продуктом азотистого обмена является аммиак. Он выделяется в воду через Жабры в виде иона аммония. Жабры проницаемы также для неорганических ионов растворенных в воде солей и низкомолекулярных соединений, транспортирование которых происходит путем диффузии или активно с помощью специальных структур.
Рис13 Микроскопическое строение Жаберного лепестка (из Yasutake, 1983)
1 — жаберный луч, 2 — лепесточки (респираторные складки), 3 — респираторный эпителии, 4 — капилляры с эритроцитами, 5 — слизистые клетки
Большую роль в осморегуляции играют хлоридные клетки, которые могут активно перемещать ионы натрия и хлора в направлении, противоположном градиенту концентрации (диффузии). V пресноводных рыб хлоридные клетки переносят одновалентные ионы из воды в кровь для замещения ионов, утраченных с обильно выделяемой мочой. У морских рыб направление переноса происходит в обратную сторону — из крови в воду.
При высоких градиентах концентраций химических веществ в воде происходит нарушение осморегуляторной функции жабр, что имеет важное значение для объяснения механизма действия многих токсикантов и используется в борьбе с болезнями рыб. Например, на этом основан гиперосмотический способ введения вакцин и лечебных препаратов.
Кожное дыхание существует у всех видов рыб, но степень участия кожи в дыхании во многом зависит от возраста рыб, условий внешней среды и других факторов. У рыб, обитающих в чистых проточных водоемах, на кожное дыхание приходится всего 3—10 %. Газообмен эмбрионов осуществляется через всю поверхность тела кожный покров и сосудистую систему желточного мешка.
Кишечное дыхание развито у вьюновых, тропических сомиков и др. Воздух, проглоченный ртом, прогоняется через кишечник и диффундирует в кровь в местах с густой сетью кровеносных сосудов.
Плавательный пузырь выполняет в основном гидростатическую функцию, но у некоторых рыб принимает активное участие вдыхании, являясь резервуаром воздуха. В зависимости от анатомии пузыря рыбы делятся на две большие группы: открытопузырные (большинство видов) и закрытопузырные (окуневые, треска, кефаль, колюшка и др.). У открытопузырных плавательный пузырь сообщается с кишечником протоком, который у закрытопузырных отсутствует. У карповых рыб плавательный пузырь делится на переднюю и заднюю камеры, которые соединены узким и коротким каналом (см. рис. 10, б).
Стенка передней камеры состоит из внутренней и наружной оболочек. Наружная оболочка в задней камере отсутствует Внутренняя выстилка обеих камер образована однослойным плоским эпителием, за которым следуют тонкий слой рыхлой соединительной ткани, мышечные тяжи и сосудистый слой. Далее расположены 2—3 эластические пластинки. Наружная оболочка передней камеры состоит из двух слоев плотной волокнистой (игольчатой) соединительной ткани, придающей ей перламутровый блеск. Снаружи обе камеры покрыты серозной оболочкой.
Наджаберный и лабиринтовый органы встречаются у змееголова и у тропических рыб (петушки, гурами, макроподы). Они представляют собой мешкообразные выпячивания жаберной полости (лабиринтовый орган) или глотки (наджаберный орган) и предназначены в основном для воздушного дыхания.
ОРГАНЫ ПИЩЕВАРЕНИЯ
Особенности строения и функционирования пищеварительной системы рыб обусловлены характером их питания и уровнем эволюционного развития. В ротовой полости рыб нет слюнных желез. Поджелудочная железа у них диффузная. Строение пищеварительного тракта мирных и хищных рыб существенно различается.
У хищников хорошо развиты зубы как на челюстях, так и на других частях полости рта (небной, сошнике, языке и др.). Но они служат лишь для захвата и удержания жертвы. У мирных рыб (сельдевых, карповых и др.) челюстные зубы отсутствуют. Для перетирания пищи у них приспособлены глоточные зубы и жерновок. В захвате и удержании пищи участвуют также жаберные тычинки.
Жаберные дуги, верхний и нижний своды рта формируют короткую глотку. За глоткой следует короткий широкий пищевод с мускулистой стенкой. Слизистая оболочка рта, глотки и пищевода выстлана многослойным плоским эпителием, содержащим многочисленные слизистые бокаловидные клетки. Подэпителиаль-ные слои образованы сетью коллагеновых волокон и поперечнополосатой мускулатурой, которая окружает весь пищевод. Наружная соединительнотканная оболочка пищевода срастается с окружающими тканями.
Желудок имеется только у хищных рвтб, а у мирных пищевод переходит непосредственно в передний отдел кишечника. Желудок чаще мешковидно расширен, хорошо растяжим и вместителен. Стенка его состоит из трех оболочек: слизистой, мышечной и серозной.
Складки слизистой высокие, покрытые призматическим эпителием. В их основании расположены трубчатые железы, открывающиеся в желудочные ямки. Под железами располагается подслизис-1&й слой, содержащий единичные мышечные пучки. Мышечная $5олочка двухслойная, хорошо развитая. Серозная оболочка обыч-jSoro строения.
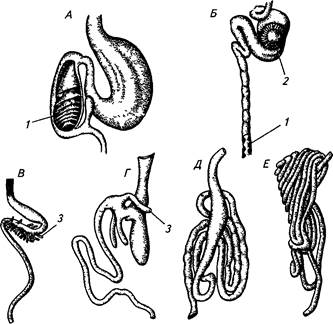
£ Кишечник обеих групп рыб не имеет четких анатомических границ и условно делится на передний, средний и задний отделы. Дли-Щ кишечника, его топография у разных видов имеют свои особенности и зависят от рода пищи. Относительная длина кишечника ра-Срггельноядных рыб составляет 6—15 (толстолобик, белый амур), Всеядных (карпа, карася) — 2—3, хищных (щуки, судака, окуня) — 0,6—1,2 (см. рис. 10, а, 14). У хищных рыб в передней части кишеч-Woca расположены слепые выросты — пилорические придатки, число которых колеблется от 3 у окуня до 400 у лососей.
Слизистая оболочка кишечника образует складки, высота кото-ВЫХ к заднему отделу постепенно уменьшается. Они покрыты каемчатым эпителием и бокаловидными клетками. К заднему отделу ки-Шечника число последних постепенно увеличивается. Подслизис-Тый слой состоит из рыхлой соединительной ткани с большим количеством лимфоидных элементов.
|
А — скат, Б-
Рис 14 Строение кишечника рыб (Моисеев, 1983) - осетр, В — лосось, Г— окунь, Д— карп, Е— толстолобик, 1 — спиральный i пан, 2— пилорическая железа, 3— пилорические придатки
ними лежат желчные и артериальные капилляры, а также синусоиды воротной вены. Триады в печени рыб встречаются редко. Секреторные трубки состоят из многоугольных клеток, содержащих круглое, бедное хроматином ядро с 1—2 крупными ядрышками.
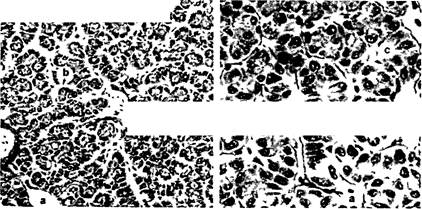
На парафиновых и целлоидиновых срезах цитоплазма гепатоцитов карповых ячеистая, содержащая много пустот, соответствующих локализации жира и гликогена. У форели и других лососевых цитоплазма гепатоцитов компактная, мелкозернистая, почти не содержащая жировых пустот. Серозная оболочка состоит из очень тонкой прослойки соединительной ткани, покрытой мезотелием (рис. 15). Печень рыб, как и других животных, является пищеварительной железой, выделяющей желчь. Она выполняет барьерную и важнейшие метаболические функции: участвует в белковом, углеводном и жировом обменах, создает запасы гликогена и жира.
Желчный пузырь овально-грушевидной формы соединен с кишечником желчным протоком, который у карповых впадает в передний отдел кишечника, а у лососевых — вблизи соединения кишки с желудком. Стенки пузыря и желчных протоков выстланы цилиндрическим эпителием, за которым следуют мышечная и серозная оболочки.
Поджелудочная железа рыб диффузная Она состоит из клеточных тяжей, сопровождающих ветви воротной вены. Она обнаруживается только гистологически. У карпа, толстолобика, белого амура, буффало и других карповых ее островки обнаруживают в мезенто-риальной клетчатке, паренхиме печени и селезенки. У лососевых она сосредоточена в мезентерии в области пилорических придатков. С кишечником железа соединяется собственным протоком, идущим рядом с желчным. Островки Ларгенганса разбросаны в
|
| «, <wv л fs-Tj. !У?дЯ? j»ed |
| - i ^* j |
Мышечная оболочка состоит из внутреннего кольцевого и наружного продольного слоев гладкой мускулатуры. Снаружи кишечник покрыт серозной оболочкой.
Печень рыб — объемистый орган, относительная масса которого у костистых достигает 8 % массы тела. Масса, плотность и цвет печени сильно колеблются в зависимости от половой зрелости рыб, времени года, интенсивности питания и качества пищи. У большинства рыб печень имеет бледно-розовую окраску с коричневатым оттенком и упругую консистенцию. Летом при интенсивном питании рыб и отложении жира в печеночных клетках она становится бледной и рыхлой. В зимний период вследствие длительного голодания печень карповых рыб часто уменьшается в объеме, становится тусклой, водянистой и нередко приобретает зеленоватую окраску за счет скопления желчного пигмента в цитоплазме гепатоцитов.
У хищных рыб печень четко обособлена, расположена в передней части брюшной полости, а у карповых находится между петлями кишечника, приобретая многолопастную форму.
Паренхима печени рыб трубчато-сетчатая, не разделяемая на дольки и состоящая из анастомозов секреторных трубок. Между
Рис 15 Микроскопическое строение печени форели (из Yasutake, 1983) а — центральные вены, Ь — синусоиды, с — печеночные клетки
скоплениях ацинарных клеток. Клеточное строение экзокринной и эндокринной частей железы не отличается от аналогичных структур теплокровных животных.
Механизм переваривания и всасывания пищи у рыб принципиально не отличается от такового у наземных животных. Желудочное пищеварение у них осуществляется с помощью пепсина и соляной кислоты, а у мирных рыб весь процесс переваривания пищи происходит в кишечнике. В нем участвуют ферменты поджелудочной железы (трипсин, химотрипсин, амилаза, мальтаза, липаза, нуклеа-зы), кишечного сока (энтерокиназа, аминопептидаза, фосфатаза и др.) и желчь. Пристеночное пищеварение и всасывание происходят в зоне щеточной каемки кишечного эпителия.
На пищеварение рыб заметно влияют факторы внешней среды: температура, химический и газовый составы воды, время года и т. д. Хотя рыбы приспособлены к длительному голоданию (до 1 года), при неблагоприятных условиях среды недостаточно упитанные и истощенные особи легко заболевают и плохо переносят воздействия различных вредных факторов.
Дата добавления: 2016-01-07; просмотров: 3044;