СЕРДЕЧНО-СОСУДИСТАЯ СИСТЕМА. КРОВЬ И ОРГАНЫ КРОВЕТВОРЕНИЯ
Главным различием кровеносной системы рыб и высших позвоночных является наличие одного круга кровообращения и двухкамерного сердца.
• Сердце расположено в сердечной полости, отделенной от брюшной полости плотной соединительнотканной перегородкой (рис. 9). Оно состоит из двух камер: тонкостенного предсердия и мускулистого желудочка. Кроме того, к ним примыкают придаточные отделы. Перед предсердием расположен венозный синус, через который венозная кровь поступает в предсердие, а из него в желудочек. К желудочку примыкает у низших рыб (хрящевые, осетровые) мышечный артериальный конус, у высших (костистых) — луковица брюшной аорты, состоящая из эластической ткани (рис. 10). Все отверстия между отделами сердца снабжены клапанами.
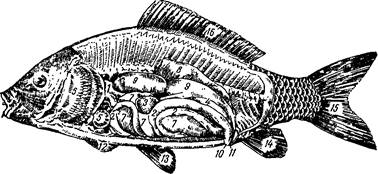
Рис. 9. Топография органов карпа:
| 1— |
— пищевод; 2 — кишечник; 3 — желчный пузырь; 4 — половые органы; 5 — сердце; 6 — жабры;
— гепатопанкреас; 8 — почки; 9 — плавательный пузырь; 10 — анальное отверстие; 11 — поло
вое отверстие; 12— плавники грудные; 13— плавники брюшные; 14—плавник анальный; /5 —
плавник хвостовой; 16— плавник спинной
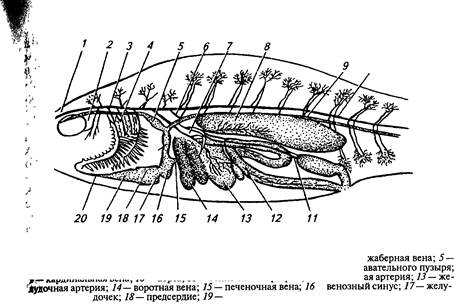
|
| 10 |
| , Рис. 11. Схема кровообращения окуня: •'Iй- обонятельная артерия; 2— лицевая артерия; 3 — головная артерия; 4— лремная вена; 6— плечевая артерия; 7— брюшная артерия; 8— артерия пл — — аая атеия 12— кишечн |
| — луковица аорты; 20— брюшная аорта |
В стенке сердца различают три оболочки: внутреннюю — эндокард, среднюю — миокард и наружную — эпикард. Наиболее мощно развит миокард желудочка, который состоит из наружного компактного и внутреннего губчатого слоев. Компактный слой образован плотным симпластом переплетающихся между собой продольных и циркулярных пучков сердечной мышечной ткани, имеющей поперечную исчерченность. Во внутреннем слое между анастомо-зирующими мышечными пучками видны небольшие полости, из-за чего он имеет губчатую структуру. Стенка предсердия и венозного синуса состоит из тонких мышечных пучков. Эндокард представлен слоем эндотелиальных клеток и тонкой прослойкой рыхлой соединительной ткани. Эпикард покрывает все отделы, состоит из слоя рыхлой клетчатки, содержащей сосуды, и снаружи покрыт мезотелием. Кровеносные сосуды (артерии, вены, капилляры и др.) у рыб имеют типичную для позвоночных структуру. Сердце у рыб иннервируется веточками блуждающего и симпатического нервов.
Рыбы имеют один замкнутый круг кровообращения (рис. 11). У костистых рыб венозная кровь из желудочка через луковицу аорты поступает в брюшную аорту, а из нее по приносящим жаберным артериям — в жабры, где насыщается кислородом. Из жабр артериальная кровь по выносящим артериям собирается в спинную аорту.
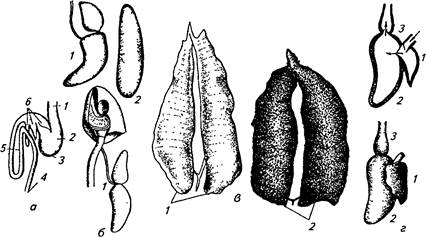
Рис. 10. Гонады, сердце, плавательный пузырь, желудочно-кишечный тракт рыб: а — кишечник хищных рыб (окунь): / — пищевод, 2 — желудок, 3 — слепой мешок, 4 — анальное отверстие, 5—средняя кишка, 6— пилорические придатки; £—плавательный пузырь: 1 — от-крытопузырных рыб (карповых), 2— закрытопузырных (окунь, судак); в —половые органы рыбы: /—семенник (сельди), 2— яичник (сельди); г — сердце: 1 — предсердие, 2—желудочек,
3 — артериальная луковица
Отпоследней к голове отходят сонные артерии, основной ствол «эрты проходит в туловищном отделе под позвоночником, а в хвостовом отделе — в гемальном канале позвоночника и называется эйюстовои артерией. От спинной аорты отходят сосуды в мускулату-•$[, кожу и во внутренние органы.
-ь Венозная кровь из хвостового отдела собирается в хвостовую «ену, которая проходит в гемальном канале позвоночника под хвос-fOBOH артерией. На уровне заднего края почек хвостовая вена разде-Швтся на две воротные вены почек, которые образуют сеть капил-АЯров, окружающую почечные канальцы. Венозные сосуды, выхо-ШПцие из почек, называются задними кардинальными венами. На чнути к сердцу они принимают вены от гонад, мускулатуры и других 'Органов. На уровне заднего отдела сердца парные задние кардинальные вены соединяются с передними кардинальными венами, собирающими кровь из головы и из области грудных плавников, и Образуют два кювьеровых протока, по которым кровь попадает в венозный синус. Кровь из пищеварительного тракта и селезенки собирается в воротную вену печени, разветвления которой в печени Образуют воротную систему. Собирающая кровь из печени печеночная вена впадает прямо в венозный синус. Следовательно, у рыб имеются две воротные системы — почек и печени. Вследствие большого разнообразия строения у разных групп рыб имеются отклонения от изложенной схемы.
Кровяное давление и скорость кровотока у рыб с удалением от сердца постепенно снижаются. У радужной форели давление в брюшной аорте составляет 40/32 мм рт. ст., в спинной аорте — 29/25 мм рт. ст. Движение крови идет за счет инерции от сокраще-
ний желудочка и скелетной мускулатуры. У взрослых рыб частота сердечных сокращений невелика — 20—45 ударов в минуту, причем у малоподвижных рыб она ниже, чем у более активных, а у молоди в несколько раз больше. У рыб последовательно расположенные сердечные камеры сокращаются поочередно, начиная с задней, т. е. первым сокращается венозный синус. Дополнительный центр регуляции ритма находится в предсердии. При повышении температуры частота сокращений увеличивается (примерно в 2—3 раза на каждые 10 °С), а при понижении замедляется. Зимой у карповых рыб частота сокращений уменьшается до 1—2 ударов в минуту, что приводит к резкому замедлению скорости кровотока, а при неблагоприятных условиях среды — к гипоксии тканей. Гипоксия, как правило, вызывает у рыб рефлекторную брадикардию, увеличение ударного объема, а также усиление гликолиза. У карпа при снижении содержания кислорода в воде до 2,5 мл/л ритм сердца остается неизменным, а при 1,8 мл/л и ниже частота сокращений сразу падает. Рецепторы, реагирующие на количество кислорода в воде, находятся в жабрах или в слизистой рта. Замедление ритма отмечено у многих рыб при извлечении их из воды, причем оно более выражено, чем при недостатке количества кислорода в воде.
Объем циркулирующей крови у рыб меньше, чем у высших позвоночных, и составляет у хрящевых ганоидов около 3,1 % массы тела, костистых — 2,2—3,6, лососевых — 3,5—4,5, у карпа — 2,0— 4,5 %(Проссер, 1978).
Морфологический и биохимический составы крови у разных видов рыб значительно различаются. Внутри вида эти показатели колеблются в зависимости от сезона, условий содержания и кормления, возраста, пола, физиологического состояния организма и т. д.
Количество эритроцитов в крови рыб меньше, чем у высших животных, а лейкоцитов, как правило, больше. Количество эритроцитов у рыб колеблется в широких пределах прежде всего в зависимости от подвижности рыб: у карпа 0,9—1,8 млн/мкл, толстолобика 1,1—2,0 млн/мкл, форели 1,2—1,8 млн/мкл, у щуки 1—2 млн/мкл. В соответствии с этим у них разный уровень гемоглобина: у карпа 7,5—10,4 г%, толстолобика 8,5—11,4 г%, форели около 10,0 г%, у щуки 7,9-9,5 г%.
Эритроциты рыб — ядерные, зрелые клетки — имеют овальную форму и ярко-оранжевую цитоплазму, незрелые — более округлую форму с разными оттенками базофилии (полихроматофилы). Причем у рыб в периферической крови эритроциты дозревают, поэтому у них встречается значительно больше незрелых эритроцитов, чем у млекопитающих.
Количество лейкоцитов у карпа колеблется от 20 до 60 тыс/мкл, у толстолобика — 60—98 тыс/мкл, форели — около 34 тыс/мкл, у щуки— 28—100 тыс/мкл. Клеточный состав лейкоцитов у рыб и высших позвоночных сходен, но резко различается по соотношению клеток. Кровь рыб имеет выраженный лимфоидный характер. В отношении классификации лейкоцитов рыб имеются противо-
чивые данные. В настоящее время принята классификация . Т. Ивановой (1983), хотя она также слишком усложнена из-за излишней детализации морфологических групп, и ее нередко трудно применять для клинического толкования изменений лейкограм-мы при патологических состояниях. Поэтому приводим более упрощенную схему (рис. I). Лейкоциты рыб делятся на агранулоциты (лимфоциты, моноциты) и гранулоциты (нейтрофилы, эозинофи-лы, базофилы).
Лимфоциты, условно делимые на большие, средние и малые, имеют типичную структуру: крупное округлое ядро, окруженное тонким слоем базофильной цитоплазмы. У рыб встречается определенное количество так называемых голоядерных форм.
Моноциты — самые крупные клетки. Ядро у них бобовидное, расположенное эксцентрично, цитоплазма дымчатая, часто вакуо-лизированная, незернистая.
Нейтрофилы — круглые клетки с овальным, палочковидным или сегментированным ядром, расположенным у края широкой зоны цитоплазмы. В зависимости от степени зрелости и формы ядра клетки делят на миелоциты, юные, палочкоядерные и сегменто-ядерные, причем у рыб немного сегментоядерных нейтрофилов. Зернистость в цитоплазме мелкая, пылевидная, окрашенная в фиолетово-розовый цвет.
Эозинофилы (псевдоэозинофилы) по морфологии сходны с нейтрофилами, но в цитоплазме имеют крупные оксифильные зерна ярко-розового цвета.
Базофилы отличаются наличием в цитоплазме базофильных зерен фиолетового цвета.
Лейкограмма разных групп и видов рыб несколько различается, но в целом имеет выраженный лимфоцитарный профиль. В ней лимфоциты составляют 80—95 %, нейтрофилы — 4—6, моноциты — 1—3 %, эозинофилы и базофилы у карпа появляются в старшем возрасте и едва достигают 1 %.
, Функции лейкоцитов рыб изучены недостаточно, но несомненно то, что они играют важную защитную роль в инфекционном процессе и, по-видимому, участвуют в детоксикации ядовитых веществ.
Тромбоциты рыб (веретеновидной формы с ядрами) обнаруживают в небольших количествах. Они участвуют в свертывании крови.
Плазма крови рыб, так же как и других животных, имеет сложный биохимический состав, выполняет трофическую и защитную функции, играет большую роль в энергетическом и пластическом обменах. Ее физико-химические показатели еще более чутко, чем морфологические, реагируют на воздействие различных внутренних и внешних факторов. Несмотря на такую лабильность, картина крови рыб, ее химический и морфологический составы в каждый данный момент отражают функциональное состояние организма, и поэтому ее исследование является важным клинико-диагностическим методом.
Органы кроветворения рыб по сравнению с высшими позвоноч-
ными животными отличаются рядом особенностей, так как у них отсутствуют костный мозг и лимфоузлы. Кроветворение у рыб происходит во многих органах, где хорошо развита ретикулярная ткань, и зависит от их систематического положения и стадии онтогенеза.
У костистых рыб во взрослом состоянии основными органами гемопоэза являются почки (головная и средняя части), селезенка и тимус. У низших рыб и в раннем онтогенезе костистых очаги кроветворения сохраняются в печени, жабрах, слизистой оболочке кишечника, под эпикардом, а иногда в стенках сосудов и других органах. Лимфоидная ткань у рыб имеет диффузное распространение по ходу кровеносных и лимфатических сосудов, а также в органах, богатых ретикулярной тканью. В случае возникновения болезни активизация лимфопоэза возможна и в этих органах. В настоящее время большинство исследователей придерживаются унитарной теории кроветворения у рыб. Исходными клетками гемопоэтической ткани являются полипотентные стволовые клетки — гемоцитобласты, которые в дальнейшем дают начало развитию всех групп клеток крови рыб, дифференцирование которых происходит по той же схеме, что и у высших позвоночных животных.
Селезенка рыб помимо депо крови является местом распада эритроцитов и кроветворным органом. Она располагается по ходу переднего отдела кишечника, имеет лентовидную форму и темно-вишневую окраску. Снаружи она покрыта капсулой из мезотелия. Пульпа содержит тонкие трабекулы и нечетко разделена на красную и белую. Строму паренхимы составляют соединительная и ретикулярная ткань, в синцитии которой расположены крупные скопления эритроцитов, а также небольшое число лимфоцитов, нейтрофилов и других клеток крови, единичные гемоцитобласты. Сосудистая сеть хорошо развита, заканчивается артериальными гильзами и венозными синусами (лакунами), из которых клетки могут выходить в пульпу селезенки. В периваскулярных зонах выявляются островки пигментных клеток, в цитоплазме которых откладываются гемосидерин и бледно-желтый пигмент типа липофусцина.
Селезенка рыб может менять свой размер не только при септических заболеваниях, но и под влиянием внешних условий и стрессовых воздействий. У карпа она увеличивается зимой, когда из-за пониженной температуры воды кровоток замедляется и кровь скапливается в селезенке, почках, печени. При воздействии стрессовых факторов (недостаток кислорода, колебания температуры, транспортирование, сортировка рыбы, лечебно-профилактические обработки и т. д.) происходит обескровливание селезенки и уменьшение ее размера, повышение кровяного давления и частоты сердечных сокращений с параллельным подъемом уровня сахара в крови и снижением количества гликогена в печени. Так, если при нормальных условиях относительная масса селезенки карпа составляет 1,2— 1,4 % массы тела, то при стрессе она уменьшается до 0,15—0,2 %, а при аэромонозе увеличивается до 1,7 % (Альбрехт, 1969).
Тимус расположен на медиальной стенке жаберной полости, вблизи места прикрепления жаберных крышек к затылочной об-дасти головы. Он состоит из соединительнотканной стромы, в которую заключены скопления лимфоцитов и видоизмененной эпителиальной ткани. Снаружи он покрыт капсулой. У многих костистых рыб железа функционирует всю жизнь, принимая участие в димфопоэзе и формировании иммунитета. Лимфатическая система у рыб представлена только сетью лимфатических щелей, капилляров и протоков. Наиболее крупные из них парные подпозвоноч-ные, головные и боковые стволы, идущие под кожей вдоль боковой линии, впадают в кардинальные вены у кювьеровых протоков. Для улучшения движения лимфы по сосудам у некоторых рыб имеются лимфатические сердца, расположенные в хвосте под пос-дедним позвонком
Дата добавления: 2016-01-07; просмотров: 2242;