ЖИЗНЕННЫЙ ЦИКЛ, РАЗМНОЖЕНИЕ
Вся жизнедеятельность рыб в течение онтогенеза определяется понятием «жизненный цикл», который делят на периоды, различающиеся рядом биологических, морфологических и физиологических особенностей.
В индивидуальном развитии рыб выделяют следующие периоды: эмбриональный, личиночный, мальковый, ювенильный, взрослого (половозрелого) организма, старческий.
Эмбриональный период начинается с момента оплодотворения яйцеклетки и продолжается до перехода молоди на питание внешней пищей.
Этот период включает в себя два подпериода: 1) собственно эмбриона, когда развитие происходит в оболочке икринки; 2) подпери-од свободного эмбриона (предличинки), когда развитие продолжается вне оболочки (после выклева); эмбрион все время питается желтком.
Личиночный период начинается с момента рассасывания желтка и перехода на питание внешней пищей. Личинки отличаются от взрослых рыб внешним и внутренним строением: у них недоразвиты жабры; нет чешуи; не дифференцированы плавники и другие органы.
Мальковый период наступает, когда организм приобретает сходство с взрослой рыбой, исчезают личиночные органы, появляются плавники, закладывается чешуя, полностью дифференцируются внутренние органы, по внешнему виду малек становится похожим на взрослых рыб.
Ювенильный (юношеский, полувзрослый) период характеризуется интенсивным ростом, развитием половых желез, появлением вторичных половых признаков (если они имеются).
Период взрослого (половозрелого) организма начинается с момента наступления половой зрелости, когда полностью сформированы половые органы, выражены вторичные половые признаки, характерные для данного вида, организм способен размножаться.
Старческий период характеризуется потерей способности размножаться, резким снижением или прекращением роста рыб.
В пределах периодов различают этапы и стадии развития рыб, знание которых важно для совершенствования биотехники разведения, понимания закономерностей развития и роста рыб, причин изменения численности популяций и т. д.
Каждый период развития имеет свое приспособительное значение и видовую специфику. В первые периоды развития до половой зрелости рыб пищевые ресурсы расходуются в основном на рост организма. В половозрелом возрасте энергетические затраты у рыб поступают преимущественно на развитие половых желез и накопление резервных веществ для поддержания обмена в период голодания, во время миграции, зимовки, размножения.
Начальным периодом жизни рыб, как и других животных, является размножение, которое у рыб отличается тем, что им свойственны наружное оплодотворение и развитие эмбрионов во внешней среде. Это влечет за собой большую гибель потомства на ранних стадиях развития в результате воздействия неблагоприятных факторов среды.
Для сохранения вида в процессе эволюции у рыб выработались защитные механизмы: большая плодовитость, забота о потомстве, живорождение и т. д.
Рыбы способны размножаться в самых разнообразных условиях. В зависимости от особенностей размножения, места откладывания икры выделяют следующие экологические группы рыб.
Литофилы — размножаются на каменистом грунте обычно в реках, на течении или на дне олиготрофных озер (осетровые, лососи, подусты и др.).
Фитофилы — размножаются среди растений, откладывая икру на вегетирующие или на отмершие растения (сазан, карп, лещ, щука, плотва, окунь и др.)- Они имеют клейкую икру.
Псаммофилы — откладывают икру на песок, иногда прикрепляя ее к корешкам растений (пелядь, ряпушка, пескарь и др.).
Пелагофилы — выметывают икру в толщу воды. Икра и свободные эмбрионы развиваются, плавая в толще воды, обычно в благоприятных кислородных условиях. В эту группу входят почти все виды сельдей, тресковых, камбал, некоторые карповые (чехонь, толстолобики, амуры и др.).
Остракофилы — откладывают икру внутрь мантийной полости моллюсков, иногда под панцири крабов (некоторые пескари, горчаки).
Наиболее плодовиты из них пелагофилы, затем следуют фито-филы и литофилы. Плодовитость рыб тесно связана с размером икринок: у рыб с крупной икрой (лососевые) она ниже, с мелкой — выше. Например, у кеты при диаметре икринок 7—8 мм плодовитость составляет 2—4 тыс. шт., у трески при диаметре икринок 1,1— 1,7мм — до 10 млн шт. Значительное влияние на плодовитость оказывают обеспеченность кормами, возраст и размер рыб. Наибольшая плодовитость отмечается при хорошей кормовой базе у крупных рыб среднего возраста. Различают плодовитость индивидуальную (абсолютную), относительную и рабочую.
Индивидуальная плодовитость — это общее количество икры, выметанное самкой за один нерестовый период (например, у 6-летнего карпа — в среднем 900 тыс. шт.).
Относительная плодовитость — количество икры, приходящееся на единицу массы самки (у карпа норма 180 тыс. шт/кг массы тела). Этот показатель нужен для сравнения плодовитости рыб в зависимости от возраста и массы: до определенного возраста он увеличивается, потом снижается.
Рабочая плодовитость — количество икры, получаемое от одной самки при проведении искусственного осеменения.
Половая зрелость у разных видов наступает в разном возрасте, причем самцы обычно созревают на год раньше самок. К наиболее скороспелым относятся рыбы с коротким жизненным циклом, которые созревают в годовалом возрасте (бычки, каспийская килька, хамса, снеток). Рыбы с длительным жизненным циклом (осетровые) становятся половозрелыми поздно (от 7—8 до 18—20 лет). Карп обычно созревает в возрасте 4—5 лет. Однако на скорость полового созревания большое влияние оказывают температура и наличие пищи. Поэтому карп в южных регионах созревает раньше, чем в северных.
У большинства рыб вторичные половые признаки развиты слабо и заметны только в период нереста, когда появляется брачный наряд (главным образом у самцов). Например, у карповых и сигов на голове, жаберных крышках, плавниках и на теле появляется жемчужная сыпь (жесткие бугорки), у хариусов краснеют плавники, у гольцов и некоторых видов лососей на теле появляются яркие пятна, у дальневосточных лососей изменяются челюсти и образуется горб (нерка, горбуша). Более четко и довольно красочно проявляются половые признаки у многих аквариумных рыб.
По срокам икрометания различают рыб:
1) нерестящихся весной (сельди, радужная форель, щука, окунь,
плотва);
2) нерестящихся летом (сазан, карп, линь, красноперка и другие
карповые);
3) нерестящихся в осенне-зимний период (многие лососи, сиги,
налим, навага).
Это деление весьма условно, так как время нереста во многом зависит от температуры, климата, а также биологии рыб, многие из которых (например, осетровые) имеют озимые и яровые формы.
По продолжительности периода икрометания выделяют рыб с единовременным и порционным нерестом. У рыб единовременного икрометания икра откладывается вся сразу и за короткое время (вобла, окунь, карп и др.). При порционном икрометании рыбы откладывают икру в несколько приемов через разные промежутки времени (несколько раз за лето). Порционность икрометания характерна для рыб тропиков и субтропиков; в умеренных широтах их меньше, а на севере почти нет. Порционное икрометание способствует увеличению плодовитости рыб за счет лучшей обеспеченности пищей и выживаемости потомства.
Большинство рыб не заботится о потомстве. Нередки случаи, когда производители (гамбузии, карпа и др.) поедают икру и особенно молодь. Поэтому их целесообразно после нереста отсаживать. В то же время многие рыбы охраняют потомство, причем эту функцию чаще выполняют самцы. Примеры заботы о потомстве самые разнообразные. Колюшки, например, строят гнезда из травы, самки тиляпии вынашивают икру и некоторое время держат личинок во рту. Лабиринтовые рыбки строят гнездо из пены, состоящей из пузырьков воздуха и слюнообразного секрета. Форель и лососи выкапывают в грунте ямки, отложенную в них икру засыпают песком и гравием. Наиболее совершенной формой заботы о потомстве является так называемое живорождение, т. е. развитие икры в половых путях самок и потом выметывание личинок в воду (гуппи, меченосцы, морской окунь и др.).
Оплодотворение яйцеклетки рыб происходит в воде путем внедрения сперматозоидов в икринки. Сперматозоиды рыб становятся подвижными только в воде и сохраняют подвижность всего 1-3 мин, потом погибают. В спермиальной жидкости они неподвижны; при хранении в холодильнике сохраняют свои оплодотворяющие свойства в течение 1—2сут. На этом основаны хранение и транспортирование «сухой» спермы. У рыб наблюдают избирательность оплодотворения. Поэтому использование при осеменении спермы двух (и более) самцов повышает оплодотворяемость икры.
После оплодотворения икринки набухают, в них происходит обводнение желтка с образованием вокруг него перивителлинового пространства, заполненного жидкостью. Перивителлиновое пространство образуется и в неоплодотворенных икринках, после чего они обычно погибают. Яйцеклетки костистых рыб относятся к те-лолецитальному типу. В них ядро и плазма клетки находятся на анимальном полюсе, а желток — на противоположной части (вегетативном полюсе). В результате дробление охватывает только верхнюю часть клетки, начинаясь с образования бластодиска. Дальше развитие идет по общей схеме: образование бластулы, морулы, гаст-рулы. При гаструляции клетки анимального полюса окружают желток с образованием двух зародышевых листков (экто- и энтодермы). Далее развитие переходит в стадию органогенеза, подвижного эмбриона, заканчиваясь выклевом предличинок. Продолжительность инкубации икры у разных видов рыб различная и зависит от температуры воды. Например, при оптимальных температурах (20— 22 °С) икра карпа развивается в течение 3-4 сут, а радужной форели при температуре около 10 °С — в течение 45—60 сут.
Свободный эмбрион (предличинка) продолжает питаться за счет желточного мешка и проходит заключительные этапы развития (формирование органов дыхания, кровообращения, пищеварения и др.). В процессе расходования желтка предличинка постепенно переходит на смешанное, а затем на экзогенное питание и превращается в личинку. Личиночный период у карпа длится около 2—3 нед, затем примерно к месячному возрасту формируются мальки.
В эмбриональном периоде отмечают переломные, уязвимые стадии, когда наблюдаются интенсивный обмен и повышенный отход эмбрионов (гаструла, органогенез).
При использовании перезревшей икры, нарушении температурного и кислородного режимов инкубации отмечаются пониженная оплодотворяемость, повышенный отход икры, появляется много уродливых личинок с укороченным туловищем, искривлением позвоночника, водянкой околосердечной и брюшной полостей и др.
ПИТАНИЕ И РОСТ РЫБ
Естественные корма рыб. При рыбохозяйственном освоении водоемов в первую очередь необходимо учитывать спектр питания рыб разных видов и возрастов, с тем чтобы максимально использовать естественные кормовые ресурсы. На этом основан принцип формирования ихтиофауны в прудах при выращивании рыб в поли-культуре, которая предусматривает смешанную посадку в пруды рыб с разным спектром питания. Знание характера и закономерностей питания имеет важное значение для кормления рыб в условиях аквакультуры, подбора необходимой рецептуры кормов и разработки техники кормления рыб, а также оценки полноценности кормления и определения причин возникновения болезней рыб.
В биоценозе естественных водоемов различные гидробионты наиболее прочно взаимосвязаны по типу питания, т. е. в них образуется трофическая (пищевая) цепь. В общем виде она состоит из трех основных звеньев:
1) водоросли, водные растения, детрит, образующие первичную
продукцию;
2) зоопланктон, зообентос, образующие промежуточную про
дукцию при потреблении организмов первого звена;
3) рыбы мирные и хищные, образующие конечную продукцию
при потреблении организмов второго и частично первого звена.
У растительноядных рыб эта трофическая цепь короткая (водоросли—рыбы), у хищников—длинная, включающая в себя все звенья.
Естественной пищей рыб являются различные водные животные и растения. Низшие беспозвоночные организмы, обитающие в толще воды, называют планктоном. Растительные организмы образуют фитопланктон, животные — зоопланктон.
Организмы, населяющие дно водоема, называют бентосом. Его подразделяют на фитобентос и зообентос.
Фитопланктон состоит из низших водорослей — диатомовых, зеленых, синезеленых и др. (рис. 17). Водоросли используют для своего роста органические и минеральные соединения, т. е. растворенные в воде соли азота, фосфора, калия, кальция, железа и др. Отмирающие водоросли служат пищей для микроорганизмов и простейших, которые развиваются на отмирающих клетках, разлагая их до более простых органических и минеральных соединений.

Рис. 17. Фитопланктон, водоросли:
| - осцилатория; б— зе- |
| а — синезеленые. , леные" 5— вольвокс, _ ,___ ,. мариум, // — анкистродесмус, 12— монас; в — диатомовые: 16 ---- |
:: 1 — анабена, 2 — афанизоменон, 3 — микроцистис, 4 — осцшштория; о — зе-;окс, 6 — хлорелла, 7— педиаструм, 8— сценодесмус, 9— пандорина, 10— кос-
---------- мус, 12— спирогира, 13 — кладофора, 14— улотрикс, 15— хламидо-
— астерионелла, 17— диатома, 18— сибелла, 19— циклотелла, 20— синедра
Зеленые водоросли имеют зеленую окраску. Клетки зеленых водорослей имеют твердую оболочку и весьма разнообразную форму. К зеленым водорослям относят вольвоксовые, протококковые, жгутиковые и др.
Синезеленые водоросли отличаются от других водорослей синезеле-ной окраской и особенностями строения. Они не имеют оформленного ядра и хроматофоров. Для них типично обильное выделение слизи, обволакивающей как отдельные клетки, так и целые колонии.
Диатомовые водоросли желто-бурого цвета характеризуются наличием кремниевого панциря, состоящего из двух створок. По форме они очень разнообразны. Встречаются водоросли в виде нитей, звездочек, а также веретенообразной формы.
При благоприятных условиях (высокая температура, обилие органических веществ и др.) водоросли развиваются в массовых количествах, вызывая так называемое цветение воды. Обычно при цветении максимально развиваются 1—2 вида водорослей. Различают основные типы цветения — протококковое, диатомовое, синезеленое и др.
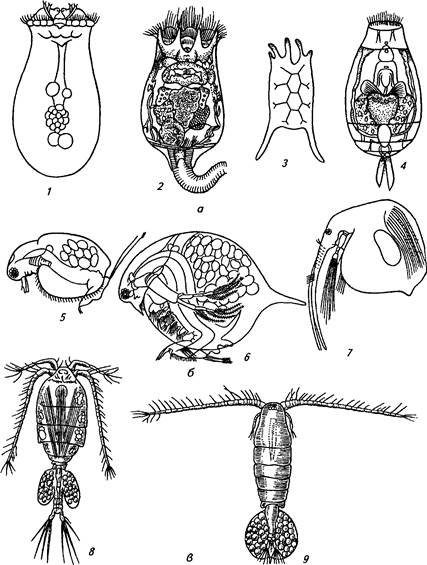
В состав зоопланктона входят инфузории, коловратки, веслоногие и ветвистоусые рачки.
Инфузории (тип простейших) представлены парамециями, инфузориями-туфельками и др.
Коловратки — мельчайшие из многоклеточных организмов — весьма разнообразны и многочисленны в пресных водоемах. Они размножаются партеногенетически. Самка, вылупившаяся из оплодотворенного яйца, на третьи сутки достигает половой зрелости. Живет примерно 2-3 нед (рис. 18, а).
Ракообразные принадлежат к числу естественных кормов для питания рыб. Низшие ракообразные представлены в пресных водах отрядами ветвистоусых Cladocera, веслоногих Copepoda и ракушко-вых Ostracoda рачков.
Ветвистоусые рачки, или кладоцеры, представляют собой одну из важнейших групп пресноводного зоопланктона. Ветвистоусые ракообразные имеют 4—7 пар ног и двуветвистые антенны. К ним относятся дафнии, цериодафнии, босмины (рис. 18, б).
Подавляющая часть кладоцер — самки, размножающиеся летом в основном партеногенетически. Развитие яиц протекает в течение нескольких суток под покровом панциря, и выводковую сумку покидают уже сформировавшиеся рачки. Один или два раза в год появляются мелкие самцы. Половое размножение обычно происходит осенью, и оплодотворенные яйца остаются на зимовку, а рачки отмирают. Главной пищей для них служат фитопланктон и бактерии. Кладоцеры занимают существенное место в питании молоди, а рыбы-планктонофаги (ряпушка, рипус, пелядь, пестрый толстолобик, веслонос и др.) питаются зоопланктоном постоянно.
Веслоногие рачки — копеподы — наряду с кладоцерами составляют существенную часть зоопланктона. Удлиненное тело веслоногих рачков подразделено на головогрудь и брюшко, оканчивающееся вилкой и хвостовыми щетинками. Веслоногие рачки размножаются только половым путем. Из яиц вылупляются личинки — наугошу-сы, имеющие три пары конечностей. Науплиусы имеют небольшие размеры (до 0,3 мм) и служат кормом для молоди рыб так же, как и взрослые формы.
В пресных водоемах веслоногие рачки представлены циклопами и диаптомусами (рис. 18, в). Циклопы — хищники. Они охотятся за простейшими, коловратками, ветвистоусыми рачками, иногда нападают на личинок рыб. Диаптомусы — фильтраторы, питаются бактериями, низшими водорослями и др. В отличие от кладоцер большинство копепод не отмирают осенью, и в зимнее время зоопланктон состоит исключительно из них.
Зообентос включает в себя организмы, относящиеся к разным систематическим группам — членистоногим, моллюскам, червям.
Из членистоногих основное население донной фауны составляют разнообразные личинки насекомых (стрекоз, поденок, веснянок, ручейников, комаров, мошек).
Наибольшее кормовое значение для карпа имеют хирономи-
|
ды — личинки комаров-толкунцов (рис. 19). Питаются личинки детритом, бактериями, растительностью. Они живут в воде 2— 3 мес, превращаются в куколок, из которых выходят взрослые комары. Заметное место занимают малощетинковые черви (олигохе-ты) — тонкие длинные черви, покрытые щетинками. Они, питаясь детритом, служат пищей для рыб. Из олигохет в прудах чаще встречается трубочник.
Моллюски в прудах представлены двумя классами: брюхоногие и двустворчатые. Из брюхоногих моллюсков наиболее распространены прудовики, катушки, живородки, битинии.

Рис. 18. Зоопланктон:
а - коловратки: / - аспланха, 2 - брахионус, 3 - керателла квадрата, 4 - лекана; 6 - ветвисто-усые рачки: J- моина, 6— дафния пулекс, 7— босмина корегони; е- веслоногие рачки- 8-
циклоп, 9—диаптомус
Рис. 19. Организмы бентоса:
хирономиды; 2— олигохеты; 3 — личинка вислокрылки; 4— шаровка; 5—прудовик; 6-бокоплав; 7— личинка комара; 8— ручейник; 9— поденка; 10— веснянка
К фитобентосу относятся представители высшей водной растительности — рогоз, камыш, тростник, осока, аир и др., которыми питаются белые амуры.
Физиология питания и рост рыб.У рыб известны два типа питания: эндогенное (за счет внутренних ресурсов организма) и экзогенное (за счет внешней пищи). Эндогенное питание свойственно эмбрионам всех видов рыб, когда они на стадии икры и предличин-ки питаются за счет желтка. Во взрослом состоянии эндогенное питание наблюдается у рыб, которые не питаются зимой (карповые), а также у проходных рыб во время нерестовых миграций (осетровые, лососи, угри, некоторые сельди). У дальневосточных лососей и угрей организм во время миграций настолько сильно истощается, что после нереста они погибают.
По характеру экзогенного питания рыб делят на две большие группы: мирных и хищных. Мирные рыбы питаются растительностью, беспозвоночными и детритом. К ним относятся планктонофа-ги (сельди, сиговые и др.), бентофаги (лещ, карп, сазан и многие другие карповые) и фитофаги, или растительноядные (толстолобик, белый амур, красноперка и др.). Хищники питаются рыбой. Однако это деление весьма условно: многие рыбы при отсутствии специфической пищи могут переходить на другие объекты питания — бентофаги потребляют зоопланктон, растительные объекты или детрит, а некоторые мирные рыбы становятся хищниками. Особенно широк спектр питания у карпа, за счет чего его относят к всеядным рыбам.
Изменение характера и интенсивности питания обусловлено рядом факторов: возрастом, полом, состоянием здоровья, временем года, химическим и температурным режимами водоема и др. Экзогенное питание рыб начинается с личиночного возраста, когда рассасывается желток. При этом следует подчеркнуть, что личинки всех видов рыб питаются одинаковой пищей: вначале поедают инфузорий, коловраток, затем переходят на ракообразных (дафний, циклопов и др.), т. е. питаются зоопланктоном. По мере роста примерно в мальковом возрасте они переходят на специфический тип питания или значительно расширяют спектр потребления кормовых объектов.
Суточное потребление пищи также зависит от возраста: молодь обычно ест больше, чем взрослые и старые рыбы. В преднерестовый период интенсивность питания снижается, а многие морские и особенно проходные рыбы питаются мало или совсем прекращают питаться. Суточная ритмика питания также различается у разных рыб. У мирных рыб, особенно планктоноядных, перерывы в питании невелики, а у хищных они могут длиться более суток. У карповых рыб отмечаются два максимума активности питания: утром и вечером.
На интенсивность питания большое влияние оказывает физиологическое состояние рыб, а также условия среды — температура, содержание кислорода и др. Упитанная рыба потребляет корма меньше, чем исхудавшая. Например, годовики карпа после зимнего голодания питаются значительно активнее, чем сеголетки в кон-
це лета. Сильно ослабляется интенсивность питания при различных заболеваниях рыб.
Из физико-химических факторов среды первостепенное значение имеют температура и газовый режим. Каждый вид рыб может питаться только в определенных температурных границах, выше и ниже которых рыба не берет пищу. Так, для карпа оптимальный температурный диапазон питания составляет 20—27 "С, у форели — 16—20 °С. При температуре ниже 4 °С и выше 35 °С карп не питается. Столь же важное значение для питания рыб имеет кислородный режим водоема. Например, при содержании кислорода менее 4 мг/л аппетит у карпа ухудшается, при 2,0 мг/л потребление корма уменьшается в 2—4 раза, а при более низком содержании кислорода вообще прекращается. Непосредственное представление о характере питания рыб дает анализ содержимого кишечника (по количеству и видовому составу кормовых объектов). Он проводится путем контрольных вскрытий рыб.
Для определения уровня потребления и усвоения пищи используют кормовой коэффициент, т. е. количество корма, необходимое для прироста единицы массы рыб. Оптимальные значения кормового коэффициента искусственных комбикормов для карпа составляют 3,5—4,5.
Другими важными показателями уровня обеспеченности рыб пищей являются коэффициент упитанности и содержание жира в теле (жирность). Коэффициент упитанности рыб определяют по формуле Фультона:
где К — коэффициент упитанности; Р— масса тела, г; / — длина тела от рыла до концг! чешуйного покрова, см.
У физиологически полноценных карпов коэффициент упитанности осенью должен составлять у сеголетков 2,7—3,4, у годовиков 1,7-2,3, у двухлетков 2,6-3,2. Этот показатель особенно важно учитывать при оценке зимостойкости рыб; его низкие значения свидетельствуют о недостаточной подготовленности карпов к зимовке. Содержание жировых запасов в организме рыб также характеризует упитанность и уровень резистентности организма. У одних видов отложение жира четко локализовано: у лососей — в мышцах, у трески — в печени, у судака — в полости тела, а у осетров, сельдей и других рыб жир распределен более равномерно по всем органам. По содержанию жира в мышцах рыб разделяют на 4 группы: тощих (0,2-1,2% жира) —щука, бычки, навага, треска, окунь, судак; среднежирных (1,5-4,5% жира) —вобла, камбала, лещ, сазан, карп; жирных (5-15 % жира) — лососи, осетровые, скумбрия; особо жирных (более 15 % жира) — угорь, минога, хамса. Содержание жира в тушке сеголетков карпа перед посадкой их на зимовку должно составлять 3-6 % их массы, что имеет важное значение для ус-
тойчивости рыб в зимний период. Запасы жира у половозрелых рыб служат источником энергии для развития гонад. Однако чрезмерное ожирение производителей отрицательно сказывается на их плодовитости и качестве потомства, а нередко приводит к бесплодию, особенно аквариумных рыб.
Рост рыб неразрывно связан с питанием и периодами развития организма. Специфической особенностью рыб является преобладание процессов ассимиляции над процессами диссимиляции, благодаря чему они растут практически всю жизнь. Но периодичность роста у них зависит от вида и возраста, а также от времени года и обеспеченности рыб кормом. В первый год жизни, особенно на стадии личинки и малька, рыбы растут очень быстро, увеличивая свою массу за сезон в несколько десятков раз. Затем рост постепенно замедляется, но продолжается и после половой зрелости. Летом, в период интенсивного питания, отмечается самая высокая скорость роста. Осенью и особенно зимой, когда температура воды снижается до 2—4 °С и ниже, теплолюбивые рыбы (карповые и др.) перестают питаться и расти. Более того, зимой у карпа отмечают потерю массы и уменьшение линейного размера. Холодолюбивые рыбы (форель, пелядь, чир, окунь, судак и др.) хотя и питаются, но их рост тоже замедляется. В связи с этим на чешуе, в плавниковых лучах и слуховых косточках образуются годичные кольца, по которым определяют возраст рыб.
В прудовом рыбоводстве и других видах аквакультуры используют зоотехнический принцип учета возраста рыб: начиная с момента рождения (весны—лета) и до половой зрелости. Для этого последовательно различают следующие возрастные группы рыб:
личинки — с момента смешанного питания до начала закладки чешуи, у карпа — примерно до 14 дней;
мальки — все тело покрыто чешуей, рыбка похожа на своих родителей, у карпа — примерно до месячного возраста;
сеголетки (0+) — вполне сформировавшиеся рыбки, прожившие лето, осень и зиму, т. е. в возрасте до 1 года;
годовики (1) — перезимовавшие сеголетки в возрасте 1 год;
двухлетки (1+) — рыбы, прожившие 1 год и одно лето, т. е. до 2 полных лет.
Далее до половой зрелости обозначение идет по тому же принципу — двухгодовики (2) -»трехлетки (2+) -> трехгодовики (3) и т. д.
При достижении половой зрелости рыб, способных размножаться, переводят в группу производителей (самцов и самок).
Продолжительность жизни рыб, их максимальные размеры и масса весьма различны и специфичны для каждого вида. Можно отметить общую закономерность: чем медленнее рыба растет и созревает, тем она дольше живет и крупнее по размеру. Например, тропические рыбы (бычки и некоторые карповые) имеют размер до 10-14 мм и живут около года, а осетровые (белуга) живут до 100 лет, достигая массы 500-1000 кг. Однако предельный возраст наших пресноводных рыб меньше: у карася 12-15 лет, укарпа, сазана, сома и других видов около 30 лет.
Дата добавления: 2016-01-07; просмотров: 4900;