Различия ферментного состава органов и тканей. Органоспецифичные ферменты. Изменения активности ферментов в процессе развития. Изоферменты и энзимодиагностика.
1.Органоспецифические ферменты:
А) секреторные (холинэстераза, церулоплазмин, про- и антико-агулянты)
Б) индикаторные (ЛДГ,АЛТ, АСТ, ГлДГ, сукцинатДГ, малатДГ, фруктозо -1- фосфатальдолаза – Ф1ФА)
В) экскреторные (лейцинаминопептидаза, 5-нуклеотидаза, ЩФ, -глюкоронидаза, -глутаматтранспептидаза)
2.Клеточноспецифические (5-нуклеотидаза, ЩФ)
49. Регуляция активности ферментов. Различные способы активации и ингибирования ферментов.
(см.42,46)
50. Эндергонические и экзергонические реакции в живой клетке. Макроэргические соединения. Дегидрирование субстратов и окисление водорода, как источник энергии для синтеза АТФ. Направление химической реакции определяется значением ΔG (свободной энергии системы). Если эта величина отр., то р-ция протекает самопроизвольно и сопровождается уменьшением свободной энергии - экзергонические р-ции (процесс окисления глюкозы дикислородом, сопровождается уменьшением энергии Гиббса) Если ΔG положительно, то реакция будет протекать только при поступлении свободной энергии извне- эндергоническими р-ции. Макроэргические соед-я – соедюсодержащие макроэргическую связь, при гидролизе которой освобождается энергия больше 30кДж. К соед.относят 1,3дифосфоглицерат, кот.обр.в гликолизе. Сукцинил-Коа, карбомоилфосфат, креатининфосфат.
51. Представление о процессах окислительного фосфорилирования. Дыхательный контроль, разобщающие агенты. Окислительное фосфолирование - образование АТФ из аденозиндифосфорной и фосфорной к-т за счет энергии, освоб-ся при ок-ии орг. в-в в живых кл. Каждая ок. р-ция в соответствии с величиной высвобождаемой энергии «обслуживается» соответствующим дыхательным переносчиком: НАДФ, НАД или ФАД. Соотв-но своим ок-восст. потенциалам эти соединения в восстановленной форме подключаются к дыхат. цепи. Разобщение процессов дыхания и окислительного фосфорилирования происходит если протоны начинают проникать через внутреннюю мембрану митохондрий. В этом случае выравнивается градиент рН и исчезает движущая сила фосфорилирования. Хим. в-ва - разобщители называются протонофорами, они способны переносить протоны через мембрану. К таковым относятся 2,4 -динитрофенол, гормоны щитовидной железы и др. В норме скорость митохондриального транспорта электронов регул-ся содержанием АДФ. Выполнение кл. ф-ций с затратой АТФ приводит к накоплению АДФ, к-й в свою очередь активирует тканевое дыхание. Т. о., клеткам свойственно реагировать на интенс-ть клет. метаболизма и поддерживать запасы АТФ на необходимом уровне. Это свойство называется дыхательным контролем.
52. Цикл трикарбоновых кислот и его биологическое значение. 53. Биологическое значение цикла трикарбоновых кислот. Цикл Кребса – общий конечный путь окисления ацетильных групп (в виде ацетил-КоА), в к-е превращ. в пр-се катаболизма большая часть орг. молекул, играющих роль «клеточного топлива»: углеводов, жирных к-т и аминок-т. Данный цикл происходит в матриксе митох. и состоит из 8 последовательных р-ций. Нач. цикл с присоед. ацетил-КоА к оксалоацетату и обр-ия цитрата. Затем лимонная кислота (шестиуглеродное соединение) путем ряда дегидрирований и двух декарбоксилирований теряет два углеродных атома и снова в цикле К. превращается в оксалоацетат (четырехуглеродное соединение), т.е. в рез-те полного оборота цикла одна молекула ацетил-КоА сгорает до СО2 и Н2О, а молекула оксалоацетата регенерируется. Ц. К. дает поток электронов высокой энергии связ. НАДН2 и ФАДН2 форме.и затем включаеются в процесс окислительного фосфорилирования, где уже снитез.АТФ и образ.вода, При окислении 1 молек. ацетил-КоА в ц. К. и системе окислительного фосфорилирования может обр-ся 12 АТФ.
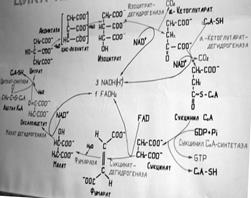 56. Амфиболические функции цикла трикарбоновых кислот.
56. Амфиболические функции цикла трикарбоновых кислот.
Дата добавления: 2015-07-22; просмотров: 4594;