Гормоны цветения. Определение пола у растений
Последующие исследования были направлены на попытку выделить гормоны цветения. Опыты показали, что длиннодневные растения зацветают на коротком дне, если их опрыснуть гиббереллином. Однако оказалось, что цветение короткодневных растений на длинном дне под влиянием опрыскивания гиббереллином не происходит. В связи с этим М.Х. Чайлахян вьщвинул гипотезу о том, что цветение вызывается двухкомпонентной гормональной системой, или одновременным наличием двух гормонов: один из этих гормонов — габбереллин, а другой — невыделенный гормон неизвестной химической природы — антезин.
Процесс зацветания растений носит двухфазный характер: на первой фазе происходит образование цветочных стеблей, а на второй — образование собственно цветков. У длнннодневных растений критической является первая фаза зацветания — образование цветочных стеблей. Именно эта фаза зависит от наличия гиббереллина, который накапливается в достаточном количестве на длинном дне. Вместе с тем у этой группы растений второй гормон цветения — антезин — всегда имеется в достаточном количестве. При выдерживании длиннодневных растений на коротком дне они не цветут, поскольку не хватает гормона гиббереллина. В связи с этим опрыскивание гиббереллином вызывает зацветание длиннодневных растений на коротком дне.
Иная картина складывается у короткодневных растений. У этой группы растений содержание гиббереллинов достаточно высокое при любой длине дня. Именно поэтому дополнительное экзогенное внесение этого фитогормона не оказывает влияния, однако у них не хватает группы фитогормонов — антезинов, которые образуются именно на коротком дне. Поскольку в листьях цветущих как коротко-, так и длиннодневных растений есть оба фитогормона, то прививка их к нецветущим растениям во всех случаях вызывает их зацветание. Показано, что цветение растений на несоответствующем фотопериоде можно вызывать, обрабатывая их экстрактом из листьев цветущих растений. У нейтральных растений зацветание также начинается при наличии двухкомпонентной гормональной системы. У этой группы растений преобладает автономная (не зависящая от внешних условий) возрастная регуляция. Биосинтез гормонов в листьях, происходящий под влиянием возрастного яровизационного или фотопериодического контроля, представляет лишь первый этап, или индукцию, цветения. Образовавшиеся гормоны перетекают к точкам роста, где и происходят дальнейшие события — преобразование вегетативной почки в цветочную, или эвокация цветения.
К сожалению, все попытки выделить антезин пока не имели успеха. Выдвигаются различные предположения о природе стимула цветения. Так, имеются данные о значении полиаминов в индукции цветения. Показано, что при падении содержания путресцина тормозится цветение горчицы белой. Еще в работах М.Х. Чайлахяна подчеркивалась роль корневой системы в индукции цветения. В последнее время получены данные, что наряду с сигналом, поступающим из листьев, в корнях образуется ряд веществ, которые транспортируются в побеги, там модифицируются и участвуют в индукции цветения. В этой связи важно, что образование цитокининов локализовано именно в корнях. Ряд исследователей полагают, что переход к цветению вызывается комплексной системой факторов, куда входят гиббереллины, цитокинины, углеводы. При этом подчеркивается роль цитокининов. Одним из первых изменений после фотопериодической индукции цветения является быстрое увеличение числа делящихся клеток. Предполагается, что в этом участвуют цитокинины.
В точках роста под влиянием притекающих туда определенных веществ и гормонов проходят изменения, перестройка апикальных меристем и переход к генеративному развитию. Одним из признаков перехода к цветению являются повышение репликации ДНК, усиление синтеза мРНК, образование специфических «репродуктивных» белков. (Э.Л. Миляева). Это может служить косвенным подтверждением, что гормоны регулируют процесс на генном уровне, в результате чего почка переходит к репродуктивному этапу морфогенеза. Показано, что у растений с терминальным типом соцветия в апексе различают центральную и медулярную зоны. Медулярная зона ответственна за рост цветочного стебля, тогда как центральная — за образование цветков. Предполагается, что гиббереллин вызывает усиление митотической активности в медулярной зоне, тогда как антезин индуцирует деление клеток в центральной зоне. Для того чтобы гормоны оказали влияние, клетки апекса должны быть восприимчивы, или компетентны. Эта компетенция может быть достигнута, по-видимому, лишь при определенном возрасте растения.
Вопрос о формировании пола у растений относится к самым сложным и наименее изученным проблемам биологии развития растений. Вместе с тем его изучение имеет непосредственное значение для разработки приемов повышения урожайности многих сельскохозяйственных культур, особенно у тех, которые образуют раздельнополые цветки. Представления о наличии разных половых форм у растений постепенно стали складываться при разработке приемов выращивания таких культур как финиковая пальма, конопля и др. Многие ученые исследовали вопросы пола у растений. В конце XVII в. Р. Камерариус установил дифференциацию пола у растений. В XVIII в. К. Линней создал представление о половой системе растений, а И. Кельрейтер экспериментально доказал существования у растений пола, В XIX в. С.Г. Навашин открыл двойное оплодотворение у покрытосеменных. В изучение проблемы пола растений крупный вклад внесли М.Х. Чайлахян, В,Н. Хрянин, Е.Г. Минина, ДА. Сабинин, Н.И. Львова, Л.И. Джапаридзе и др.
Под определением (детерминацией) пола у растений понимается формирование признаков пола у клеток, органов или особей под воздействием, как генетических факторов, так и условий внешней и внутренней среды. По наличию и степени развития генеративных органов цветки делят на обоеполые (гермафродитные) и однополые (раздельнополые). Последние бывают пестичными (женскими) или тычиночными (мужскими). На одном растении могут находиться цветки разных типов. Если цветки формируются на одних и тех же организмах, то такие растения называют однодомными. Двудомными являются растения, имеющие на одних экземплярах пестичные цветки, а на других — тычиночные (хмель, конопля, тополь). Растения с гермафродитными цветками составляют более 70% видов, только 4—5% видов растений двудомны.
У ряда двудомных растений (элодея, некоторые мхи) были обнаружены различия между половыми хромосомами. Однако эти различия выражены менее четко, чем у животных организмов. Обнаружение растений (70 видов) геретохромосом типа XX и XV позволило исследовать основы генетического определения пола у растений. Установлено, что гены, ответственные за детерминацию пола у растений располагаются не только в половых хромосомах, но и в других хромосомах (аутосомах).
Следующий за определением пола процесс его формирования происходит у растений в разные периоды онтогенеза и определяется в основном влиянием внешних условий. Дело в том, что в отличие от животных организмов процесс формирования пола у растений подвержен большим колебаниям в онтогенезе растений. Под действием различных причин может происходить даже полное превращение одного пола в другой. Это связано с регуляторными процессами, обусловленными действием факторов внешней среды, и внутренними изменениями метаболического характера.
Многочисленные факты свидетельствуют о влиянии на проявление признаков пола у растений таких внешних воздействий как влажность почвы и воздуха, температура, спектральный состав света, условия минерального питания (рисунок). Так, появлению женских цветков и формированию женских растений у двудомных видов (женская сексуализация) способствуют низкие температуры, высокая влажность, хорошее азотное питание. Дифференцировке мужских цветков (мужская сексуализация) — высокие температуры, низкая влажность, калийное питание.
Изучение явления фотопериодизма привело к выяснению, что характер дифференциации пола зависит от длины дня. Показано, что короткий день приводит
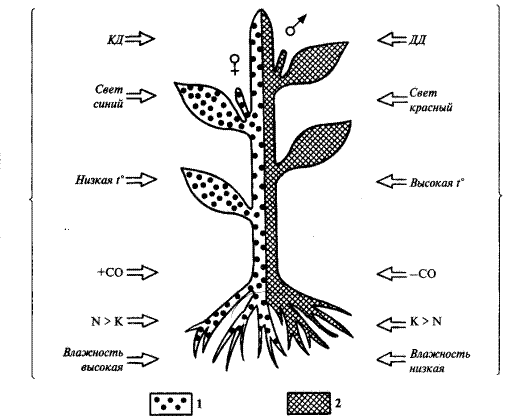
Рисунок - Роль факторов внешней среды и фитогормонов в проявлении пола у растений
Дата добавления: 2015-06-05; просмотров: 2359;