Регуляция процессов развития
Процесс развития — переход от одного этапа в другой — обусловливается внутренними причинами, заложенными в генетической основе организма. В геноме каждого организма зашифрована общая программа развития, которая делится на ряд подпрограмм. Однако должны существовать особые переключатели, которые переводят развитие организма с одной подпрограммы на другую. Иначе говоря, должны быть какие-то индукторы, регулирующие процесс экспрессии генов, т. е. вызывающие репрессию одних генов, вырабатывающих белки-ферменты для осуществления предыдущего этапа, и одновременно — дерепрессию других генов, ответственных за развертывание процессов метаболизма, необходимых для прохождения последующего этапа. Такими индукторами, по-видимому, являются гормоны.
Таким образом, последовательность развертывания программы и сами подпрограммы строго детерминированы в наследственной основе. Однако продолжительность отдельных этапов, время наступления следующего этапа могут заметно варьироваться в зависимости от условий среды. Именно под влиянием определенного комплекса внешних условий возникает такое соотношение гормонов, которое вызывает дерепрессию определенных участков генома. Как следствие, образуются белки-ферменты, изменяется течение обмена веществ, происходят морфогенетические изменения, характерные для следующего этапа. Для развертывания программы и перехода к следующему этапу развития недостаточно наличия гормонов. Клетка должна быть восприимчива (компетентна) к гормональному воздействию.
Влияние внешних условий на процесс развитияЦентральным моментом развития растительного организма является переход от вегетативного роста к репродуктивному развитию (у цветковых растений — к цветению). Условия среды могут влиять на реализацию генетической информации и тем самым ускорять или замедлять наступление определенных этапов развития, в первую очередь переход растения от этапа молодости к этапу зрелости.
Согласно представлениям, развиваемым в работах М.Х. Чайлахяна, в растительном организме существуют два типа регуляторных механизмов, которые влияют на переход растения от этапа молодости к зрелости,— автономный и индуцированный. Автономная регуляция — это возрастной контроль, не зависящий от условий среды. Автономность надо понимать как возможность перехода к цветению вне зависимости от внешних условий. Индуцированная регуляция вызывается определенными факторами среды.
Автономная регуляция проявляется в том, что растение лишь в определенном возрасте переходит в цветочно-спелое состояние. Период, который требуется для того, чтобы достигнуть цветочно-спелого состояния, различается в зависимости от вида растений. Существуют растения, у которых он почти отсутствует. Так, некоторые растения готовы при определенных условиях перейти к цветению уже на фазе семядольных листьев. Однако у большинства растений цветочно-спелое состояние наступает при наличии нескольких листьев. У многолетних растений для перехода в это состояние обычно требуется пять или более лет. Некоторые виды бамбука вступают в цветочно-спелое состояние лишь по прошествии 50 лет.
Различные исследователи по-разному объясняют физиологическую сущность этого явления. Некоторые придают значение образованию нужного количества и качества питательных веществ (трофический фактор). Так, Г. Клебс (1905) придавал решающее значение для перехода растений к цветению соотношению углерода к азоту. Клебс полагал, что все условия, вызывающие накопление Сахаров, способствуют зацветанию растений, тогда как условия, ведущие к накоплению азотистых веществ, поддерживают вегетативное состояние. Таким образом, переход к цветению связывался с накоплением определенных трофических веществ. Действительно, для некоторых растений это подтвердилось. Так, у плодовых деревьев с помощью кольцевания можно ускорить образование цветочных почек.
По данным М.Х. Чайлахяна, правило Клебса оказалось верным только для длиннодневных растений. Большое значение в переходе в цветочно-спелое состояние имеет накопление гормональных веществ. Благодаря возрастной автономной регуляции в растениях возникает градиент цветочно-спелого состояния. Это выражается в определенной последовательности зацветания побегов разных ярусов. При этом опыты показали, что цветочно-спелое состояние зависит от накопления определенных веществ, по-видимому, гормонального типа, образующихся в коре стеблевых сегментов, в узлах определенного яруса. Однако для многих видов растений достижение цветочно-спелого состояния зависит не только от возраста, но и от внешних условий.
Для индуцированной, зависящей от внешних воздействий регуляции особенное значение имеет воздействие двух факторов — температуры и фотопериода. Зависимость перехода растений к цветению от температуры называют яровизацией; зависимость перехода растений к цветению от соотношения длины дня и ночи в течение суток — фотопериодизмом. Оба эти явления выработались в процессе эволюции как приспособления, обеспечивающие переход к цветению в наиболее благоприятное время. С помощью этих приспособлений растительный организм определяет время цветения.
Яровизация. Для ряда растений переход к этапу зрелости (образованию цветочных зачатков) происходит лишь после воздействия в течение определенного периода пониженными температурами. Впервые это было подмечено Г. Гасснером (1918). При этом все растения можно разделить на три группы: 1) растения, которые не переходят к репродукции без воздействия в течение определенного времени пониженными температурами,— это главным образом озимые формы, двулетники и некоторые многолетники; 2) растения, у которых развитие ускоряется при воздействии пониженными температурами, однако это воздействие не является обязательным (салат, шпинат и др.); 3) растения, которые не требуют для перехода к цветению пониженных температур.
Особенно ясно зависимость развития от температуры (яровизация) проявляется на двулетних растениях, у ряда растений ранневесенней флоры, а также на озимых формах. Многие двулетние растения (свекла, капуста, сельдерей и др.) в первый год жизни остаются в вегетативном состоянии и переходят к репродукции лишь после воздействия пониженных температур в зимний период. Известно, что при весеннем высеве озимые формы (рожь, пшеница, ячмень) интенсивно растут, кустятся, но, в отличие от яровых форм, не переходят к репродукции, не выколашиваются. Различия между озимыми и яровыми формами зерновых культур обусловлены генетически.
Исследования ряда ученых показали, что для перехода к репродукции озимые растения нуждаются в течение определенного времени в воздействии пониженных температур. В естественных условиях при осеннем высеве озимые растения подвергаются длительному влиянию пониженных температур и нормально выколашиваются. Именно эта особенность озимых растений позволяет им переносить зимние условия. Оказалось, что, если подвергнуть действию пониженной температуры наклюнувшиеся семена озимых культур в течение определенного времени, они переходят к репродукции (выколашиванию) и при весеннем высеве. Именно это и дало основание для введения термина «яровизация». Необходимо отметить, что яровизация не вызывает непосредственно перехода растений к репродукции, а лишь подготавливает этот переход. Иначе говоря, для того чтобы в растении появились цветочные зачатки, необходимы определенные физиолого-биохимические изменения, которые проходят у некоторых видов растений под воздействием пониженных температур.
Яровизационные изменения происходят в точках роста. Для того чтобы прошли яровизационные изменения, именно точка роста должна быть подвергнута действию пониженных температур. Оказалось, что достаточно выдержать изолированные зародыши при пониженных температурах, чтобы выросшие из них растения оказались подготовленными к цветению. Клетки, образовавшиеся из точек роста, подвергшихся действию пониженных температур, уже являются яровизированными. Если меристематические клетки находились при пониженных температурах и в них произошли соответствующие изменения в обмене веществ, то эти изменения передаются при делении к вновь появляющимся клеткам и, наконец, обусловливают переход растения от вегетативного роста к репродукции. Растения могут проходить яровизацию в разном возрасте, начиная с наклюнувшихся семян. Однако существуют растения (некоторые двулетние), у которых действие низких температур воспринимается апексами при наличии листьев. Яровизация может быть эффективна даже во время развития зародыша на материнском растении, если после оплодотворения прошло не более 5 дней.
Пониженные температуры, которые необходимы разным растениям для прохождения яровизационных изменений, несколько различаются. Наиболее эффективными для озимой пшеницы являются температуры от 4 до 0°С. Длительность воздействия также колеблется. Для большинства сортов озимой пшеницы длительность воздействия пониженными температурами должна составлять 35—60 суток. Такой длительный срок показывает, что происходящие под влиянием пониженных температур изменения накапливаются постепенно. Для прохождения яровизационных изменений необходимо достаточное снабжение водой и кислородом. Последнее указывает на связь происходящих изменений с окислительно-восстановительными процессами, в частности с дыханием.
Показано, что под воздействием пониженных температур в меристематических клетках изоэлектрическая точка белков сдвигается в кислую сторону. Имеются сведения, что при яровизации происходят изменения в нуклеиновом обмене. Заметно увеличивается количество РНК. Однако в целом сущность биохимических изменений, происходящих под влиянием пониженных температур и приводящих к тому, что в растении может осуществляться переход к репродукции и развертываться соответствующая подпрограмма, не представляется ясной. Некоторые исследователи (А. Ланг) высказывают предположение, что существует особый, способный к передвижению фактор (верналин), вырабатывающийся при пониженных температурах и вызывающий переход к репродукции у требующих яровизации растений.
Возникает вопрос, каким образом реакции, возникающие при яровизации, могут ускоряться при понижении температуры. Предполагается, что яровизация зависит от двух реакций. Одна из них (б) имеет более высокий, а другая (а) более низкий температурный коэффициент (Q10). Обе реакции имеют общее исходное вещество, которое при повышении температуры превращается в неактивный, а при понижении — в активный продукт согласно схеме:
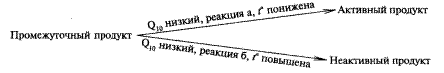
В этом случае при понижении температуры более интенсивно будет идти реакция (а) и активный продукт накапливается. Яровизационные изменения обратимы. Под воздействием повышенных температур возможна разъяровизация, в результате чего готовность клеток к образованию репродуктивных органов теряется. В некоторых случаях действие пониженных температур может быть заменено выращиванием растений при непрерывном освещении.
Дата добавления: 2015-06-05; просмотров: 2324;