ВВЕДЕНИЕ. ИСТОРИЯ ФИЗИОЛОГИИ 4 страница
Дифференцировочное торможение вырабатывается при подкреплении одного условного сигнала (например, звук с частотой 500 Гц) и отсутствии подкрепления сходный с ним сигналов (звук 1000,200 и 100 Гц), на которые первоначально (в период генерализации условного рефлекса) получался условный ответ. Этот вид торможения, вчастности, позволяет спортсмену отдифференцировать сокращения ненужных мышц при выработке двигательного навыка, т. е. имеет важное координационное значение. Процесс воспитания человека сопровождается постоянной дифференцировкой подкрепляемых и осуждаемых обществом поведенческих реакций (что такое «хорошо» и что такое «плохо»).
Запаздывающее торможение формируется при отставлении на определенный отрезок времени подкрепления от условного сигнала. В этом случае сразу после условного сигнала реакция отсутствует (Тормозится), но перед моментом подкрепления обнаруживается.
В жизни обычно встречаются не отдельные условные рефлексы, а сложные их комплексы, в которых они сочетаются с безусловными рефлексами (двигательными, сердечно-сосудистыми, дыхательными и пр.). Систему условных и безусловных рефлексов И. П. Павловназвал динамическим стереотипом. Она вырабатывается при повторении одного и того же порядка раздражений (ситуаций) и, соответственно, выражается в цепи закрепленных ответных реакций, т. е. стереотипе. Но при этом изменение внешних условий может вызвать перестройку этой системы или ее разрушение, что отмечается термином — динамический. Например, у собаки выработан динамический стереотип на определенный порядок из 6 раздражителей, и имеются на них закрепленные условные величины слюноотделения, специфические для каждого сигнала: 1)Свет— 12 капель; 2) Звук —20 к.; 3) Метроном-120 уд. • с1 — 10 к.; 4) Метроном —60уд. • с1 (неподкрепляемый раздражитель) — 0 к.; 5) Свет — 12 к.; 6) Звук—20 к. Если теперь подавать один и тот же сигнал, то ответная цепь реакций сохранится прежней: 1)Свет— 12к.;2)Свет —20к.;3)Свет— 10к.;4)Свет —Ок.; 5) Свет — 12 к.; 6) Свет — 20 к. Однако изолированное включение светового раздражения сохраняет обычный ответ —12 к. Следовательно, в коре больших полушарий собаки образована цепьпоследо- вательно возбуждающихся или затормаживающихся нервных центров, в которой активность каждого автоматически вызывает включение следующего. Подобный стереотип возникает у спортсмена при выработке двигательного навыка, особенно при выполнении стандартных движений. Такой стереотип, связанный с цепью моторных актов, А. Н. Крестовников назвал двигательный динамический стереотип. Он легче образуется при выполнении циклических упражнений, чем ациклических.
' 4.4. ТИПЫ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ,
I И II СИГНАЛЬНАЯ СИСТЕМА
Случившееся в 1924 г. в Ленинграде сильное наводнение грозило затопить клетки с подопытными собаками, которые испытали сильный стресс. На следующий день обнаружилось, что у некоторых из них пропали прочно выработанные условные рефлексы, но удругих рефлексы сохранились. Это навело И. П. Павлова на мысль о различных типах нервной системы у животных. В качестве основных свойств нервной системы И. П. Павлов рассматривал силу возбуждения и торможения, их уравновешенность и подвижность. С учетом этих свойств им были выделены следующие 4типа высшей нервной деятельности (ВИД), которые оказались сходными с 4темперамен- тами, выделенными еще Гиппократом в V веке до н. э.
1.Тип сильный неуравновешенный (холерик). Характеризуется сильным процессом возбуждения и более слабым процессом торможения, поэтому легко возбуждается и струдом затормаживаетсвои реакции. '
2.Тип сильный уравновешенный и высокоподвижный (сангвиник). Отличается сильными уравновешенными и высокоподвижными процессами возбуждения и торможения. Легко переключается с одной формы деятельности на другую, быстро адаптируется к новой ситуации.
3.Тип сильный уравновешенный инертный (флегматик). Имеет сильные и уравновешенные процессы возбуждения и торможения, но мало подвижный — медленно переключающийся с возбуждения на торможение и обратно. С трудом переходит от одного вида дея - тельности кдругому, зато вынослив при длительной работе. Медленно, но прочно адаптируется к необычным условиям внешней среды.
4.Тип слабый (меланхолик). Характеризуется слабыми процессами возбуждения и торможения, с некоторым преобладанием тормозного Процесса, мало адаптивен, подвержен неврозам. Зато обладает высокой чувствительностью к слабым раздражениям и может их легко дифференцировать.
Описанныетипы имеются уживотныхи человека. Они представляют собой лишь крайние проявления особенностей нервной системы, между которыми может быть значительное число переходных типов.
Кроме того, И. П. Павлов выделил специфически человеческие типыВНД, связанные с наличием у человека особой — второй сигнальной системы— слова видимого, слышимого, написанного, произносимого, в отличие от первой сигнальной системы, общей для человека и животных — непосредственных раздражителей внешней или внутренней среды организма. Вторая сигнальная система чрезвычайно расширила адаптационные возможности человека. Ее свойствами являются — обобщение стпзлоъ I и 11 сигнальной системы, появление абстракций (сложных комплексных понятий — мужество, ярость, доброта и пр.), возможность передачи накопленного опыта предшествующих поколений последующим (возникновение науки, культуры и пр.). Вторая сигнальная система таким образом составила основу письменной и устной речи, появления математических и нотных символов, абстрактного мышления человека. Ее деятельность связывают с функциями третичных полей коры больших полушарий, преимущественно левого полушария у правшей, где находятся центры речи.
В связи с различным соотношением улюдей реакций, связанных с преобладанием I или II сигнальной системы, И. П. Павлов различал специфически человеческие типы нервной системы: «мыслительный» — с преобладанием второй сигнальной системы—и *художе- ственный» — с преобладанием первой сигнальной системы. Среди взрослыхлюдей количество лиц с преобладанием второй сигнальной системы составляет около половины населения. Около 25% составляют лица с преобладанием первой сигнальной системы и примерно 25% — лица, имеющие равновесие обеих систем. Соответственно этим типам, в настоящее время различают 2основные формы интеллекта человека: невербальный интеллект, отражающий природные возможности индивида манипулировать с непосредственными (особенно зрительно-пространственными) раздражителями, и вербальный интеллект, отражающий способность манипулировать со словесным материалом, что определяет характер поведенческих реакций, в том числе и в спорте.
5. НЕРВНО-МЫШЕЧНЫЙ АППАРАТ
У человека существует 3 вида мышц: поперечно-полосатаые скелетные мышцы, особая поперечно-полосатая сердечная мышца и гладкие мышцы внутренних органов.
5.1. ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ СКЕЛЕТНЫХ МЫШЦ
Скелетные мышцы человека содержат около 300 млн. мышечных волокон и имеют площадь порядка 3 м2. Целая мышца представляет собой отдельный орган, а мышечное волокно — клетку. Мышцы иннервируются двигательными нервами, передающими из центров моторные команды, чувствительными нервами, несущими в центры информацию о напряжении и движении мышц, и симпатическими нервными волокнами, влияющими на обменные процессы в мышце. Функции скелетных мышц заключаются в перемещении частей тела друг относительно друга, перемещении тела в пространстве (локомоция) и поддержании позы тела.
Функциональной единицей мышцы является двигательная единица, состоящая из мотонейрона спинного мозга, его аксона (двигательного нерва) с многочисленными окончаниями и иннервируем ых им мышечных волокон. Возбуждение мотонейрона вызывает одновременное сокращение всех входящих в эту единицу мышечных волокон. Двигательные единицы (Д Е) небольших мышц содержат малое число мышечных волокон (ДЕ мышц глазного яблока 3-6 волокон, мышц пальцев руки 10-25 волокон), а Д Е крупных мышц туловища и конечностей — до нескольких тысяч (например, ДЕ икроножной мышцы человека — около 2000 мышечных волокон).
Мелкие мышцы иннервируются из одного сегмента спинного мозга, а крутые мышцы—мотонейронами 2-3 спинальных сегментов. М о- тонейроны, иннервирующие одну мышцу, составляют общий мо- тонейронный пул, в котором могут находиться мотонейроны различных размеров. Большие ДЕ образованы крупными мотонейронами, которые имеют толстые аксоны, множество концевых разветвлений и большое число связанных с ними мышечных волокон. Такие ДЕ имеют низкую возбудимость, генерируют высокую частоту нервных импульсов (порядка 20-50 импульсов в 1 с) и характеризуются высокой скоростью проведения возбуждения. Они включаютсявработулишьпривысокихнагрузкахнамышцу. Мелки е ДЕ имеют мотонейроны небольших размеров, тонкие и медленно проводящие аксоны, малое число мышечных волокон. Они легко возбудимы и включаются в работу при незначительных мышечных усилиях. Нарастание нагрузки вызывает активацию разл ич- ных Д Е скелетной мышцы в соответствии с их размерами —■ от меньших к большим (правило Хеннемана).
Мышечное волокно представляет собой вытянутую клетку (ее диаметр около 10-100мкм, а длина 10-12 см). В состав волокна входят его оболочка — сарколемма, жидкое содержимое — саркошаз- ма,ядро, энергетические центры—митохондрии, белковые депо т- рибосомы, сократительные элементы—миофибриллы, а также замкнутая система продольных трубочек и цистерн, расположенных вдоль миофибрилл и содержащих ионы Са2*,— саркоплазматический ретикулум. Поверхностная мембрана клетки через равные проме^ жутки образует поперечные трубочки, входящие внутрь мышечного волокна, по которым внутрь клетки проникает потенциал действия при ее возбуждении.
Миофибриллы — это тонкие волокна (диаметр их 1-2мкм, длина 2-2.5 мкм), содержащие 2 вида сократительных белков (прагю* фибрилл): тонкие нити актина и вдвое более толстые нити миозина. Они расположены таким образом, что вокруг миозиновых нитей находится 6 актиновых нитей, в вокруг каждой актиновой — 3 миозиновых. Миофибриллы разделены Z-мембранами на отдельные участки— саркомеры, в средней части которых расположены преимущественно миозиновые нити, а актиновые нити прикреплены к Z-мембранам по бокам саркомера. (Разная способность актина и миозина преломлять свет создает в состоянии покоя мышцы ее поперечно-полосатый вид в световом микроскопе).
Нити актина составляют около 20% сухого веса миофибрилл. Актин состоит из двух форм белка: 1) глобулярной формы — в виде сферических молекул и 2). палочковидных молекул тропомиозина, скрученных в виде двунитчатых спиралей в длинную цепь. Напротя- жении этой двойной актиновой нити каждый виток содержит по 14
молекул глобулярного актина (по 7 молекул с обеих сторон), наподобие нитки с бусинками, а также центры связывания ионов Са 2+. В этих центрах содержится особый белок (тропонин), участвующий в образовании связи актина с миозином.
Миозин составлен из уложенных параллельно белковых нитей (эта часть представляет собой так называемый легкий меромио- зин). На обоих концах его имеются отходящие в стороны шейки с утолщениями — головками (эта часть—тяжелый меромиозин), благодаря которым образуются поперечные мостики между миозином и актином.
5.2. МЕХАНИЗМЫ СОКРАЩЕНИЯ И РАССЛАБЛЕНИЯ МЫШЕЧНОГО ВОЛОКНА
При произвольной внутренней команде сокращение мышцы человека начинается примерно через 0.05 с (50 мс). 3а это время моторная команда передается от коры больших полушарий к мотонейронам спинного мозга и по двигательным волокнам к мышце. Подойдя к мышце, процесс возбуждения должен с помощью медиатора преодолеть нервно-мышечный синапс, что занимает примерно 0.5 мс. Медиатором здесь является ацетилхолин, который содержится в синаптических пузырьках в пресинаптической части синапса. Нервный импульс вызывает перемещение синаптических пузырьков кпреси- иаптической мембране, их опорожнение и выход медиатора в синаптическую щель. Действие ацетилхолина на постсинаптическую мембрану чрезвычайно кратковременно, после чего он разрушается аце- тилхолинэстеразой на уксусную кислоту и холин. По мере расходования запасы ацетилхолина постоянно пополняются путем его синтезирования в пресинаптической мембране. Однако, при очень частой и длительной импульсации мотонейрона расход ацетилхолина превышает его пополнение, а также снижается чувствительность постсинаптической мембраны к его действию, в результате чего нарушается проведение возбуждения через нервно-мышечный синапс. Эти процессы лежат в основе периферическихмеханизмовутомления прцдлительной и тяжелой мышечной работе.
Выделившийся в синаптическую щель медиатор прикрепляется к рецепторам постсинаптической мембраны и вызывает в ней явления деполяризации. Небольшое подпороговое раздражение вызывает лшйьместное возбуждение небольшой амплитуды — потенциал концевой пластинки (ПКП).
При достаточной частоте нервных импульсов ПКП достигает порогового значения и на мышечной мембране развивается мышечный потенциал действия. Он (со скоростью 5 м • с-1) распространяется вдоль по поверхности мышечного волокна и заходите попе
речные трубочки внутрь волокна. Повышая проницаемость клеточных мембран, потенциал действия вызывает выход из цистерн и трубочек саркоплазматическогоретикулума ионов Са2+, которые проникают в миофибриллы, к центрам связывания этих Ионов на молекулах актина.
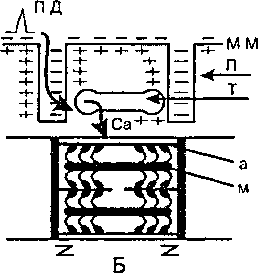
Под влиянием Са2+длинные молекулы тропомиозина проворачиваются вдоль оси и скрываются в желобки между сферическими молекулами актина, открывая участки прикрепления головок миозина к актину. Тем самым между актином и миозином образуются так называемые поперечные мостики. При этом головки миозина совершают гребковые движения, обеспечивая скольжение нитей актина вдоль нитей миозина с обоих концов саркомера к его центру, т. е. механическую реакцию мышечного волокна (рис. 10).
|
Энергия гребкового движения одного мостика производит перемещение на 1% длины актиновой нити. Для дальнейшего скольжения сократительных белков друг относительно друга мостики между актином и миозином должны распадаться и вновь образовываться на следующем центре связывания Са2*-. Такой процесс происходит в результате активации в этот момент молекул миозина. Миозин приобретает свойства фермента АТФ-азы, который вызывает распад АТФ. Выделившаяся при распаде АТФ энергия приводит к разрушению
| 1 + 11 + | ч- ч- | z--------------------------------- | ч- ч- |
| _ | Ч- ч- ч- ч- | ----- СаЗ | ч- ч~ ч- ч- |
_1HIL
z A Z
Рис. 10. Схема электромеханической связи в мышечном волокне На А: состояние покоя, на Б— возбуждение и сокращение Пд — потенциал действия, мм — мембрана мышечного волокна, п — поперечные трубочки, т — продольные трубочки и цистерны </ ионами Са2*, а — тонкие нити актина, м -- толстые нити миозина s с утолщениями (головками) на концах. Зет-мембранами ограничены« саркомеры миофибрилл, Толстые стрелки — распространение потенциала действия при возбуждении волокна и перемещение ионов Са2* из цистерн и продольных трубочек в миофибриллы, где они содействуют образованию мостиков Между нитями актином и миозином и скольжение этих нитей (сокращение волокна) за счет гребковых движений головок миозина.
имеющихся мостиков и образованию в присутствии Са2+новых мостиков на следующем участке актиновой нити. В результате повторения подобныхпроцессов многократного образования и распада мостиков сокращается длина отдельных саркомеров и всего мышечного волокна в целом. Максимальная концентрация кальция в миофиб- рилле достигается уже через 3 мс после появления потенциала действия в поперечныхтрубочках, а максимальное напряжение мышечного волокна—через 20 мс.
Весь процесс от появления мышечного потенциала действия до сокращения мышечного волокна называется электромеханической связью (или электромеханическим сопряжением). В результате сокращения мышечного волокна актин и миозин более равномерно распределяются внутри саркомера, и исчезает видимая под микроскопом поперечная исчерченность мышцы.
Расслабление мышечного волокна связано с работой особого механизма — «кальциевого насоса», который обеспечивает откачку ионов Са2+ из миофибрилл обратно в трубочки саркоплазматическо- го ретикулума. На это также тратится энергия АТФ.
5.3. ОДИНОЧНОЕ И ТЕГАНИЧЕСКОЕ СОКРАЩЕНИЕ.
ЭЛЕКТРОМИОГРАММА
При единичном надпороговом раздражении двигательного нерва или самой мышцы возбуждение мышечного волокна сопровождается одиночным сокращением. Этаформа механической реакции состоит из 3 фаз: латентного или скрытого периода, фазы сокращения и фазы расслабления. Самой короткой фазой является скрытый период, когда в мышце происходит электромеханическая передача. Фаза расслабления обычно в 1.5-2 раза более продолжительна, чем фаза сокращения, а при утомлении затягивается на значительное время.
Если интервалы между нервными импульсами короче, чем дли- тельность одиночного сокращения, то возникает явление суперпозиции — наложение механических эффектов мышечного волокна друг на друга и наблюдается сложная форма сокращения — тетанус. Различают 2 формы тетануса — зубчатый тетанус, возникающий при более редком раздражении, когда происходит попадание каждого следующего нервного импульса в фазу расслабления отдельных одиночных сокращений, и сплошной или гладкий тетанус, возникающий при более частом раздражении, когда каждый следующий импульс попадает в фазу сокращения (рис. 11). Таким образом, (в некоторых границах) междучастотой импульсов возбуждения и амплитудой сокращения волокон ДЕ существует определенное соотношение: при небольшой частоте (например, 5-8 имп. в 1с)
| f |
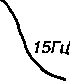
|
| г |

|
Puc. 11. Одиночное сокращение, зубчатый и сплошной тетанус камбаловидной мыищы человека (по: Зимкин Н.В. и др., 1984). Верхняя кривая — сокращение мышцы, нижняя — отметка раздражения мышцы, справа указана частота раздражения
возникают одиночные сокращения, при увеличении частоты (15-20 имп. в 1с)—зубчатый тетанус, при дальнейшем нщэастании частоты (25-60 имп. в 1 с) — гладкий тетанус. ОдиночнЬе сокращение — более слабое и менее утомительное, чем тетаническое. Зато тетанус обеспечивает в несколько раз более мощное, хотя и кратковременное сокращение мышечного волокна.
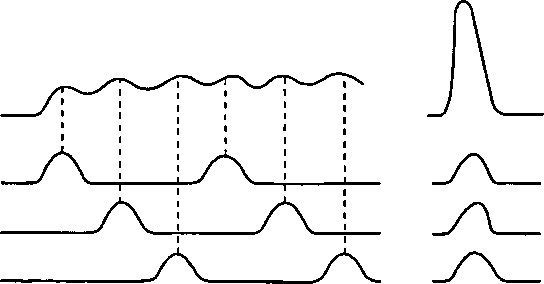
Сокращение целой мышцы зависит от формы сокращения отдельных ДЕи их координации во времени. При обеспечении длительной, но не очень интенсивной работы, отдельные ДЕ сокращаются попеременно (рис. 12), под держивая общее напряжение мышцы на заданном уровне (например, при беге на длинные и сверх- длинные дистанции). При этом отдельные ДЕ могут развивать как одиночные, так и тетанические сокращения, что зависит от частоты нервных импульсов. Утомление в этом случае развивается медленно, так как, работая по очереди, ДЕ в промежутках между активацией успевают восстанавливаться. Однако для мощного кратковременного усилия (например, поднятия штанги) требуется синхронизация активности отдельных ДЕ, т. е. одновременное возбуждение практик чески всех ДЕ. Это, в свою очередь, требует одновременной актива-
|
| Рис. 12. Различные режимы работы двигательных единиц (ДЕ) |
| Целая мышца ДЕ-1 ДЕ-2 ДЕ-3 |
ции соответствующих нервных центров и достигается в результате длительной тренировки. При этом осуществляется мощное и весьма утомительное тетаническое сокращение.
Амплитуда сокращения одиночного волокна не зависит от силы надпорогового раздражения (закон «Все или ничего»). В отличие от этого, при нарастании силы надпорогового раздражения сокращение целой мышцы постепенно растет до максимальной амплитуды.
Работа мышцы с небольшой нагрузкой сопровождается редкой частотой нервных импульсов и вовлечением небольшого числа ДЕ. В этих условиях, накладывая отводящие электроды на кожу над мышцей и используя усилительную аппаратуру, можно на экране осциллографа ил и с применением чернильной записи на бумаге зарегистрировать одиночные потенциалы действия отдельных Д Е. В случае же значительных напряжений потенциалы действия многих ДЕ алгебраически суммируются и возникает сложная интегрированная кривая записи электрической активности целой мышцы— электромиограмма (ЭМГ).
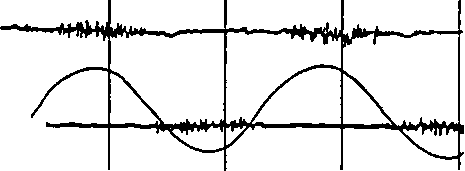
Форма ЭМГ отражает характер работы мышцы: при статических усилиях она имеет непрерывный вид, а при динамической работе — вид отдельных пачек импульсов, приуроченных, в основном, к начальному моменту сокращения мышцы и разделенных периодами «электрического молчания». Особенно хорошо ритмичность появления подобныхпачекнаблюдаетсяуспортсменовпри циклической работе (рис. 13). У маленьких детей и неадаптированных ктакой работе лиц четких периодов отдыха не наблюдается, что отражает недостаточное расслабление мышечных волокон работающей мышцы.
Чем больше внешняя нагрузка и сила сокращения мышцы, тем выше амплитуда ее ЭМГ. Это связано с увеличением частоты нервных импульсов, вовлечением большего числа ДЕ в мышце и синхронизацией
|
| ЭМГ трехгл. мышцы Механограмма |
| Рис. 13. Электромиограмма мышц-антагонистов при циклической роботе |
их активности. Современная многоканальная аппаратура позволяет производить одновременную регистрацию ЭМ Г многих мышц на разных каналах. При выполнении спортсменом сложныхдвижений можно видеть на полученных ЭМГ кривых не только характер активности отдельных мышц, но и оценить моменты и порядок их включения или выключения в различные фазы двигательных актов. Записи ЭМГ, полученные в естественных условиях двигательной деятельности, можно передавать к регистрирующей аппаратуре по телефону ил и радиотелеметрически, Анализ частоты, амплитуды и формы ЭМ Г (например, с помощью специальных компьютерных программ) позволяет получить важную информацию об особенностяхтехники выполняемого спортивного упражнения и степени ее освоения обследуемым спортсменом.
По мере развития утомления при той же величине мышечного усилия амплитуда ЭМГ нарастает. Это связано с тем, что снижение сократительной способности утомленныхгДЕ компенсируется нервными центрами вовлечением в работу дополнительных ДЕ, т, е. путем увеличения количества активных мышечных волокон. Кроме того, усиливается синхронизация активности ДЕ, что также повышает амплитуду суммарной ЭМГ.
5.4. МОРФОФУНКЦИОНАЛЬНЫЕ ОСНОВЫ МЫШЕЧНОЙ СИЛЫ
Движение является результатом взаимодействия внутренних и внешних сил, развиваемых в опорно-двигательном аппарате, — активных (возникающих при сокращении или напряжении мышцы во время ее возбуждения) и пассивных (упругое напряжение при растяжении мышцы, сопротивление мышцы и ее сухожилия).
Сила мышцы зависит отряда морфологических и физиологических фактор/щ количества и свойств мышечных волокон в мыш-
це, исходной длины мышцы, характера нервных импульсов, механических условий действия мышцы на кости скелета.
Сила мышцы является суммой силы отдельных ее мышечных волокон. Подсчитано, что 1 одиночное мышечное волокно икроножной мышцы развиваетнапряжение 100-200мг, 1 ДЕ икроножной мышцы человека содержит около 2000 мышечных волокон и развивает напряжение 200-400 г, 1 икроножная мышца содержит около 1000 ДЕ,т. е. развиваетнапряжение 200-400 кг.
Большое значение имеет анатомическое строение мышцы. В параллельно-волокнистых и веретенообразных мышцах (камбаловидная мышца и др.) сила мышц тем больше, чем больше ее анатомический поперечник, т. е. площадь поперечного сечения целой мышцы. В перистых мышцах (двуглавая мышца и др.) физиологический поперечник, т. е. площадь поперечного сечения всех мышечных волокон гораздо больше, чем ее анатомический поперечник. В такой мышце упаковано значительно больше мышечных волокон и, соответственно, больше ее сила.
На силу сокращения мышцы влияет ее исходная длина, так как от нее зависит возможное количество поперечных мостиков между актином и миозином. Предполагают, что в каждом цикле при- соединения-отсоединения поперечных мостиков расходуется энергия 1 молекулы АТФ на 1 поперечный мостик. Следовательно, чем больше образуется в мышечном волокне актино-миозиновых мостиков, тем выше скорость расщепления АТФ, больше тяга сократительных белков и, соответственно, больше развиваемая мышцей сила.
Наибольшее количество актино-миозиновых контактов образуется при небольшом растяжении мышцы до некоторой оптимальной длины. При значительном растяжении саркомера нити актина далеко расходятся в стороны и практически не контактируют с расположенным в средней части саркомера миозином. В случае же резкого уменьшения длины саркомера нити актина в центре перекрывают друг друга, препятствуя контактам с миозином и также уменьшая число образуемых мостиков. В связи с этими особенностями взаимодействия сократительных белков наибольшая сила мышцы проявляется при некотором ее предварительном растяжении.
Одной из важнейших характеристик скелетных мышц, влияющих на силу сокращения, является состав (композиция) мышечных волокон. Различают 3 типа мышечных волокон — медленные неутомляемые (I типа), быстрые неутомляемые или промежуточные (I I-атипа) и быстрые утомляемые (Н-бтипа).
Медленные волокна (1 типа), их обозначают также SO — Slow Oxydative (англ. — медленные окислительные) — это выносливые (неутомляемые) и легко возбудимые волокна, с богатым кровоснабжением, большим количеством митохондрий, запасов миоглобинаи с использованием окислительных процессов энергообразования (аэробные). Их, в среднем, у человека 50%. Они легко включаются в работу при малейших напряжениях мышц, очень выносливы, но не обладают достаточной силой. Чаще всего они используются при поддержании ненагрузочной статической работы, например, при сохранении позы.
Быстрые утомляемые волокна (11-б типа) или FG — Fast Glicolitic (быстрые гликолитические) используют анаэробные процессы энергообразования (гликолиз). Они менее возбудимы, включаются при больших нагрузках и обеспечивают быстрые и мощные сокращения мышц. Зато эти волокна быстро утомляются. Их примерно 30%. Волокна промежуточного типа (11-а) — быстрые неутомляемые, окис-< лительные, их около 20%. В среднем, для разных мышц характерно различное соотношение медленных неутомляемых и быстрых утомляемых волокон. Так, в трехглавой мышце плеча преобладают быстрые волокна (67%) над медленными (33%), что обеспечивает скоростно-силовые возможности этой мышцы (рис. 14), а д ля более медленной и выносливой камбаловидной мышцы характерно наличие 84% медленных и всего 16% быстрых волокон (Салтин Б., 1979).
Однако, состав мышечных волокон в одной и той же мыище имеет огромные индивидуальные различия, зависящие от врожденных типологических особенностей человека. К моменту рождения человека его мышцы содержат лишь медленные волокна, но под влиянием нервной регуляции устанавливается в ходе онтогенеза генетически заданное индивидуальное соотношение мышечных волокон разного типа. По мере перехода от зрелого возраста к пожилому число быстрых волокон у человека заметно снижается и, соответственно, уменье шается мышечная сила. Например, наибольшее количество быстрых волокон в наружной головке 4-х главой мышцы бедра мужчины (около 59-63%) отмечается в возрасте 20-40 лет, а в возрасте 60-65 лет их число почти на 1/3 меньше (45%).
Трехглавая Наружная Камбало-
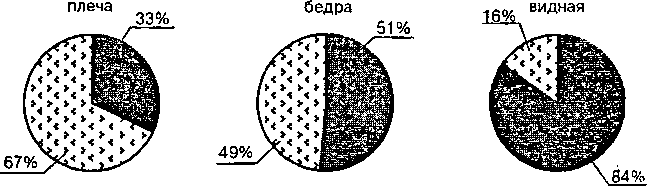 Рис. 14. Состав мышечных волокон в разных мышцах Медленные — черным цветом; быстрые — серым
Рис. 14. Состав мышечных волокон в разных мышцах Медленные — черным цветом; быстрые — серым
|
Количество тех или других мышечных волокон не изменяется в процессе тренировки. Возможно только нарастание толщины (гипертрофия) отдельных волокон, а также некоторое изменение свойств промежуточных волокон. При направленности тренировочного процесса на развитие силы происходит нарастание объема быстрых волокон, что и обеспечивает повышение силы тренируемых мышц.
Характер нервных импульсов изменяет силу сокращения мышц тремя способами: 1) увеличением числа активных ДЕ — это механизм вовлечения или рекрутирования ДЕ (сначала происходит вовлечение медленных и более возбудимых ДЕ, затем — высокопороговых быстрых ДЕ); 2) увеличением частоты нервных импульсов, в результате чего происходит переход от слабых одиночных сокращений ксильным тетаническим сокращениям мышечных волокон; 3) увеличением синхронизации ДЕ, при этом происходит увеличение силы сокращения целой мышцы за счет одновременной тягивсехак- тивных мышечных волокон.
Дата добавления: 2015-05-21; просмотров: 789;