ВВЕДЕНИЕ. ИСТОРИЯ ФИЗИОЛОГИИ 2 страница
Эфферентные нейроны (центробежные) связаны с передачей нисходящих влияний от вышележащих этажей нервной системы к нижележащим или из ЦНС к рабочим органам. Для эфферентных нейронов характерны разветвленная сеть коротких отростков —дендритов и один длинный отросток—аксон.
Промежуточные нейроны (интернейроны, или вставочные)—это, как правило, более мелкие клетки, осуществляющие связь между различными (в частности, афферентными и эфферентными) нейронами. Они передают нервные влияния в горизонтальном направлении (например, в пределах одного сегмента спинного мозга) и в вертикальном (например, из одного сегмента спинного дюзга в другие—■ выше или нижележащие сегменты). Благодаря многочисленным разветвлениям аксона промежуточные нейроны могут одновременно возбуждать большое число других нейронов.
3.2.3. ВОЗБУЖДАЮЩИЕ И ТОРМОЗЯЩИЕ СИНАПСЫ
Взаимодействие нейронов между собой (и с эффекторными орга- нами) происходит через специальные образования — синапсы (греч.—контакт). Они образуются концевыми разветвлениями нейрона на теле или отростках другого нейрона. Чем больше синапсов на нервной клетке, тем больше она воспринимает различных раздражений и, следовательно, шире сфера влияний на ее деятельность и возможность участия в разнообразных реакциях организма. Особенно много синапсов в высших отделах нервной системы иименноу нейронов с наиболее сложными функциями.
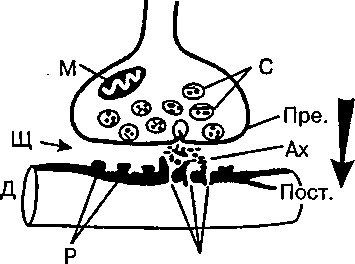
В структуре синапса различаюттри элемента (рис. 2):
1) пресинаптическую мембрану, образованную утолщением мембраны конечной веточки аксона;
2) синаптическую щель между нейронами;
3) постсинаптическую мембрану — утолщение прилегающей поверхности следующего нейрона.
Рис. 2. Схема синапса Пре. — пресинаптическая мембрана,
| Поры |
|
Пост. — постсинаптическая мембрана,
С — синаптические пузырьки, Щ — синаптическая щель,
М — митохондрии.
Ах — ацетилхолин Р — рецепторы U порь! (Порыf дендрита (Д) следующего нейрона,
Стрелка — одностороннее проведение возбуждения.
В большинстве случаев передача влияния одного нейрона на другой осуществляется химическим путем. В пресинаптической части контакта имеются синаптические пузырьки, которые содержат специальные вещества — медиаторы или посредники. И ми могут быть ацетилхолин (в некоторых клетках спинного мозга, в вегетативных узлах), норадреналин (в окончаниях симпатических нервных волокон, в гипоталамусе), некоторые аминокислоты и др. Приходящие в окончания аксона нервные импульсы вызывают опорожнение синаптических пузырьков и выведение медиатора в синаптическую щел£>.
По характеру воздействия на последующую нервную клетку различают возбуждающие и тормозящие синапсы. '
Ввозбуждающих синапсах медиаторы (например, ацетилхолин) связываются со специфическими макромолекулами по- стсинаптической мембраны и вызывают ее деполяризацию. При этом регистрируется небольшое и кратковременное (около 1 мс) колебание мембранногоПотенциалавсторонудей<мя/шда<ш!Или возбуждаю- щий постсинаптический потенциал (ВПСП).Для возбуждения нейрона необходимо, чтобы ВПСП достиг порогового уровня. Для этого величинадеполяризационного сдвига мембранного потенциала должна составлять не менее 10 мВ. Действие медиатора очень кратковременно (1 -2 мс), после чего он расщепляется на неэффективные компоненты (например, ацетилхолин расщепляется ферментом холинэстеразойнахашниуксуспую кислоту) или поглощается обратно пресинаптическими окончаниями (например, норадреналин).
В тормозящих синапсах содержатся тормозные медиаторы (например, гамма-аминомасляная кислота). Их действие на постсинаптическую мембрану вызывает усиление выхода ионов калия из клетки и увеличение поляризации мембраны. При этом регистрируется кратковременное колебание мембранного потенциала в сторону гиперполяризации—тормозящий постсинаптический потенциал (ТПСП). В результате нервная
клетка оказывается заторможенной. Возбудить ее труднее, чем в исходном состоянии. Для этого понадобится более сильное раздражение, чтобыдостичь критического уровня деполяризации.
3.2.4. ВОЗНИКНОВЕНИЕ ИМПУЛЬСНОГО ОТВЕТА НЕЙРОНА
На мергбране тела и дендратов нервной клетки находятся как возбуждающие, так и тормозящие синапсы. В отдельные моменты времени часть их может быть неактивной, а другая часть оказывает активное влияние на прилегающие к ним участки мембраны. Общее изменение мембранного потенциала нейрона является результатом сложного взаимодействия (интеграции) местных ВПСП и ТПСП всех многочисленных активированных синапсов. При одновременном влиянии как возбуждающих, так и тормозящих синапсов происходит алгебраическое суммирование (т.е. взаимное вычитание) их эффектов. При этом возбуждение нейрона возникнет лишь в том случае, если сумма возбуждающихпостсинаптическихпотенциалов окажется больше суммы тормозящих. Это превышение должно составлять определенную пороговую величину (около 10 мВ).Тольков этом случае появляется потенциал действия клетки. Следует отметить, что в целом возбудимость нейрона зависит от его размеров: чем меньше клетка, тем выше ее возбудимость.
С появлением потенциала действия начинается процесс проведения нервного импульса по аксону и передача его на следующий нейрон или рабочий орган, т.е. осуществляется эффекторная функция нейрона. Нервный импульс является основным средством связи между нейронами.
Таким образом, передача информации в нервной систем происходит с помощью двух механизмов — электрического (ВПСП; ТПСП; потенциал действия) и химического (медиаторы).
3.3. ОСОБЕННОСТИ ДЕЯТЕЛЬНОСТИ НЕРВНЫХ ЦЕНТРОВ
Свойства нервных центров в значительной мере связаны с особенностями проведения нервных импульсов через синапсы, связывающие различные нервные клетки.
3.3.1. ОСОБЕННОСТИ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ЧЕРЕЗ НЕРВНЫЕ ЦЕНТРЫ
Нервным центром называют совокупность нервных клеток, необходимых для осуществления какой-либо функции. Эти центры отвечают соответствующими рефлекторными реакциями на внешнее
раздражение, поступившее от связанных с ними рецепторов. Клетки нервных центров реагируют и на непосредственное их раздражение веществами, находящимися в протекающей через них крови (гуморальные влияния). В целостном организме имеется строгое согласование — координация их деятельности.
Проведение волны возбуждения от одного нейрона к другому через синапс происходит в большинстве нервных клеток химическим путем — с помощью медиатора, а медиатор содержится лишь в преси- наптической части синапса и отсутствует в постсинаптической мембране. Поэтом '(важной особенностью проведения возбуждения через синаптические контакты является одностороннее проведение нервных влияний, которое возможно лишь от пресинапти- ческой мембраны к постсинаптической и невозможно в обратном направленииЗв связи с этим поток нервных импульсов в рефлекторной дуге имеет определенное направление от афферентных нейронов к вставочным и затем к эфферентным — мотонейронам или вегетативным нейронам.
Большое Значение в деятельности нервной системы Имеет другая особенность проведения возбуждения через синапсы — замедленное проведение. Затрата времени на процессы, происходящие от момента подхода нервного импульса к преси- наптической мембране до появления в постсинаптической мембране потенциалов,называется синаптической задержкой. В большинстве центральных нейронов она составляет около
0.3 мс. После этого требуется еще время на развитие Возбуждающего постсинаптического потенциала (ВПСП) и потенциала действия. Весь процесс передачи нервного импульса (от потенциала действия одной клетки до потенциала действия следующей клетки) через один синапс занимает примерно 1.5 мс. При утомлении, охлаждении и ряде других воздействий длительность синаптической задержки возрастает. Если же для осуществления какой-либо реакции требуется участие большого числа нейронов (многих сотен и даже тысяч), то суммарная величина задержки проведения по нервным центрам может составить десятые доли секунды и даже целые секунды.
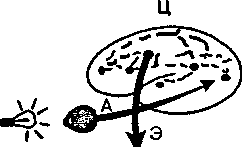
При рефлекторной деятельности общее время отмомента нанесения внешнегораздражения до появления ответной реакции организма—так называемое скрытое или латентное время рефлекс af определяется в основном длительностью проведения через синапсы. Величина латентного времени рефлекса служит важным показателем функционального состояния нервных центров. Измерение латентного времени простой двигательной реакции человека на внешний сигнал широко используется в практике для оценки функционального состояния ЦНС (рис. 3).
Рис. 3. Схема измерения времени двигательной реакции А — афферентные,
|
Э — эфферентные и Ц — центральные пути; С— отметка светового сигнала,
| t 150мс |
О — отметка нажима кнопки,
1150мс — время реакции.
3.3.2. СУММАЦИЯ ВОЗБУЖДЕНИЯ
В ответ на одиночную афферентную волну, идущую от рецепторов кнейронам, в пресинаптической части синапса освобождается небольшое количество медиатора. При этом в постсинаптической мембране нейрона обычно возникает ВПСП — небольшая местная деполяризация. Для того, чтобы общая по всей мембране нейрона величина ВПСП достигала порога возникновения потенциаладействия, требуется суммацияна мембране клетки многих подпороговых ВПСП. Лишь в результате такой суммации возбуждения возникает ответ нейрона. Различают пространственную и временную суммацию.
Пространственная суммация наблюдается в случае одновременного поступления нескольких импульсов в один и тот же нейрон по разным пресинаптическим волокнам. Одномоментное возбуждение синапсов в различных участках мембраны нейрона повышает амплитуду суммарного ВПСП до пороговой величины. В результате возникает ответный импульс нейрона и осуществляется рефлекторная реакция. Например, для получения ответа двигательной клетки спинного мозга обычно требуется одновременная активация 50-100 афферентных волокон от соответствующих периферических рецепторов..
Временная суммация происходит при актйвации одного и того же афферентного пути серией последовательных раздражений. Если интервалы между поступающими импульсами достаточно коротки и ВПСП нейрона от предыдущих раздражений не успевают затухать, то последующие ВПСП накладываются друг на друга, пока деполяризация мембраны нейрона не достигнет критического уровня для возникновения потенциала действия)\Таким способом даже слабые раздражения через i (екоторое время могут вызывать ответные реакции организма (например, чихание и кашель в ответ на слабые раздражения слизистой оболочки дыхательных путей).
3.3.3. ТРАНСФОРМАЦИЯ И УСВОЕНИЕ РИТМА
Характер ответного разряда нейрона зависит не только от свойств раздражителя, но и от функционального состояния самого нейрона (его мембранного заряда, возбудимости, лабильности). Нервные клетки обладают свойством изменять частоту передающихся импульсов, т. е, свойством трансформации ритма.
При высокой возбудимости нейрона (например, после приема кофеина) может возникать учащение импульсации (мультипликация ритма), а при низкой возбудимости (например, при утомлении) происходит урежете ритма, так как несколько приходящих импульсов должны суммироваться, чтобы наконец достичь порога возникновения потенциаладействия. Эти изменения частоты импульсации могут усиливать или ослаблять ответные реакции организма на внешние раздражения.
При ритмических раздражениях активность нейрона можетна- строиться на ритм приходящих импульсов,т. е. наблюдается явление усвоения ритма (Ухтомский А. А., 1928). Развитие усвоения ритма обеспечивает сонастройку активности многих нервных центровпри управлении сложными двигательными актами, осб- бенноэто важно для поддержания темпа циклических упражнений.
3.3.4. СЛЕДОВЫЕ ПРОЦЕССЫ
После окончания действия раздражителя активное состояние нервной клетки или нервного центра обычно продолжается еще некоторое время. Длительность следовых процессов различна: небольшая в спинном мозге (несколько секунд или минут), значительно больше в центрах головного мозга (десятки минут, чась1 или даже дни) и очень большая в коре больших полушарий (до нескольких десятков лет).
Поддерживатьявное и кратковременное состояние возбуждения в нервном центре могут импульсы, циркулирующие по замкнутым цепям нейронов. Значительно сложнее по природе длительно сохраняющиеся скрытые следы. Предполагают, что длительное сохранение в нервной клетке следов со всеми характерными свойствами раздражителя основано на изменении структуры составляющих клетку белков и на перестройке синаптических контактов.
Непродолжительные импульсные последействйя (длительностью до 1 часа) лежат в основе так называемой кратковременной памяти, адлительные следы, связанные со структурными и биохимическими перестройками в клетках, — в основе формирования долговременной памяти.
Процессы координации деятельности ЦНС основаны на согласовании двух нервных процессов — возбуждения и торможения. Торможение является активным нервным процессом, который предупреждает или угнетает возбуждение.
3.4.1. ЗНАЧЕНИЕ ПРОЦЕССА ТОРМОЖЕНИЯ В ЦНС
Явление торможения в нервных центрах было впервые открыто И* М, Сеченовым в 1862 г. Значение этого процесса было рассмотрено им в книге «Рефлексы головного мозга» (1863).
Опуская лапку лягушки в кислоту и одновременно раздражая некоторые участки головного мозга (например, накладывая кристаллик поваренной соли на область промежуточного мозга), И. М. Сеченов наблюдал резкую задержку и даже полное отсутствие «кислотного» рефлекса спинного мозга (отдергивания лапки). Отсюда он сделал заключение, что одни нервные центры могут существенно изменять рефлекторную деятельность в других центрах, в частности вышележащие нервные центры могут тормозить деятельность нижележащих. Описанный опыт вошел в историю физиологии под названием Сеченовское торможение. >
Тормозные процессы — необходимый компонент в координации нервной деятельности. Во-первых, процесс торможения ограничивает распространение возбуждения на соседние нервные центры, чем способствует его концентрации в необходимых участках нервной системы. Во-вторых, возникая в одних нервных центрах параллельно с возбуждением других нервных центров, процесс торможения тем самым вьиаиочает деятельность ненужных в данныймамент органов. В-треть- их, развитие торможения в нервных центрах предохраняет их от чрезмерного перенапряжения при работе, т. е. играет охранительную роль.
3.4.2. ПОСТСИНАПТИЧЕСКОЕ И ПРЕСИНАЛТИЧЕСКОЕ ТОРМОЖЕНИЕ
Процесс торможения, в отличие от возбуждения, неможетрас- нространяться по нервному волокну—это всегда местный процесс в области синаптических контактов. По месту возникновения различают пресинаптическое и постсинаптическое торможение.
Постсинаптическое торможение — это тормозные эффекты, возникающие в постсинаптической мембране. Чаще всего этот вид торможения связан с наличие^ в ЦНС специальных тормозных нейронов. Они представляют собой особый тип вставочных нейронов, у которых окончания аксонов выделяют тор- мозный медиатор. Одним изтаких медиаторов является гамма-аминомасляная кислота (ГАМ К).
Нервные импульсы, подходя к тормозным нейронам, вызыёают в них такой же процесс возбуждения, как и в других нервных клетках. В ответ по аксону тормозной клетки распространяется Ьбычный потенциал действия. Однако, в отличие от других нейронов, окончания аксона при этом выделяют не возбуждающий, а тормозной медиатор. В результате тормозные клетки тормозят те нейроны, на которых оканчиваются их аксоны.
К специальным тормозным нейронам относятся клетки Рэншоу в спинном мозге, клетки Пуркинье мозжечка, корзинчатЫе клетки h промежуточном мозге идр. Большое значение, например, тормозные клетки имеют при регуляции деятельности мышц-антагонистов: приводя к расслаблению мышц антагонистов, они облегчаюттем Самым одновременное сокращение мышц-агонистов (рис.4).
Клетки Рэншоу участвуют в регуляции уровня активности отдельных мотонейронов спинного мозга. При возбуждении мотонейрона импульсы поступают по его аксону кмышечным волокнйм и одновременно по коллатералям аксона—ктормозной клетке Рэн- шоу. Аксоны последней «возвращаются» к этому же нейрону, вызывая его торможение. Чем больше возбуждающих импульсов посылает мотонейрон на периферию (а значит, и ктормозной клетке), Тем сильнее это возвратное торможение (разновидность постсинаптического торможения). Такая замкнутая система дей-
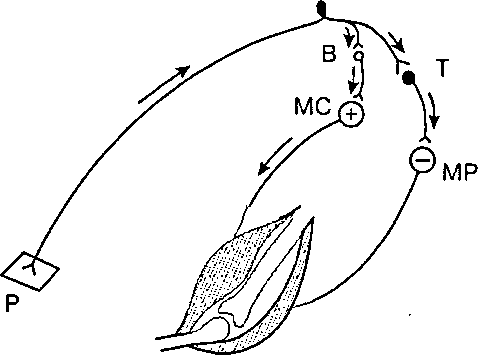 Рис. 4. Участие тормозной клетки в регуляции мышц-антагонистов ВиТ— возбуждающий и тормозный нейроны. Возбуждение (+) Мотонейрона мышцй-сгибателя (МС) и торможение (—) мотонейрона мышцы - разгибателя (МР). Р — кожный рецептор.
Рис. 4. Участие тормозной клетки в регуляции мышц-антагонистов ВиТ— возбуждающий и тормозный нейроны. Возбуждение (+) Мотонейрона мышцй-сгибателя (МС) и торможение (—) мотонейрона мышцы - разгибателя (МР). Р — кожный рецептор.
|
ствует какмеханизм саморегуляции нейрона, предохраняя его от чрезмерной активности. •
Клетки Пуркинье мозжечка своими тормозными влияниями на клетки подкорковых ядер и стволовых структур уча- ствуютв регуляции тонуса мышц.
КорзиНчатые клетки в промежуточном мозге являются как бы воротами, которые пропускают или не пропускают импульсы, идущие в кору больших полушарий от различных областей тела.
Пресинаптическое торможение возникает перед синаптическим контактом—В пресинаптической области. Окончание аксона тормозной нервной клетки образует синапс на конце аксона возбуждающей нервной клетки, вызывают чрезмерно сильную деполяризацию мембраны этого аксона, которая угнетает проходящие здесь потенциалы действия и тем самым блокирует передачу возбуждения. Этот вид торможения ограничивает поток афферентных импульсов к нервным центрам, выключая посторонние для основной деятельности влияния.
3.4.3. ЯВЛЕНИЯ ИРРАДИАЦИИ И КОНЦЕНТРАЦИИ
При раздражении одного рецептора возбуждение может в принципе распространяться в ЦНСвлюбомнаправлении и налюбуюнервную клетку. Это происходит благодаря многочисленным взаимосвязям нейронов одной рефлекторной дуги с нейронами других рефлекторных дуг. Распространение процесса возбуждения на другие нервные центры называют явлением иррадиации.
Чем сильнее афферентное раздражение и чем выше возбудимость окружающих нейронов, тем больше нейронов охватывает процесс иррадиации. Процессы торможения ограничивают иррадиацию и способствуют концентрации возбуждения в исходном пункте ЦНС.
Процесс иррадиации играет важную положительную роль при формировании новых реакций организма (ориентировочных реакций, условных рефлексов). Чем больше активируется различных нервных центров, тем легче отобрать из их числа наиболее нужные для последующей деятельности центры. Благодаря иррадиации возбуждения между различными нервными центрами возникают новые функциднальные взаимосвязи—условные рефлексы. Н а этой основе возможно, например, формирование новых двигательных навыков.
Вместе с тем, иррадиация возбуждекия может оказать и о/п/ш- цателъное воздействие на состояние и поведение организма, нарушая тонкие взаимоотношения между возбужденными и заторможенными нервными центрами и вызывая нарушения координации движений.
3.4.4. ДОМИНАНТА
Исследуя особенности межцентральных отношений, А, А. Ухтомский обнаружил, что если в организме животного осуществляется сложная рефлекторная реакция, например, повторяющиеся акты глотания, то электрическое раздражение моторных центрцв не только перестает вызывать в этот момент движение конечностей, но и усиливает протекание начавшейся цепной реакции глотания, которая оказалась главенствующей.
Такой господствующий очаг возбуждения в ЦНС, определяющий текущую деятельность организма, А, А. Ухтомский (1923) обозначил термином доминанта.
Доминирующий очаг может возникнуть при повышенном уровне возбудимости нервных клеток, который создается различными гуморальными и нервным^ влияниями. Он подавляет деятельность других центров, оказывая сопряженное торможение,
Объединение большого числа нейронов в одну доминантную систему происходит путем взаимного сонастраивания на общий темп активности, т. е. путем усвоения ритма. Одни нервные клетки снижают свой более высокий темп деятельности, а другие — повышают низкий темп до некоторого среднего, оптимального ритма. Доминанта может надолго сохраняться в скрытом, следовом состоянии (потенциальная доминанта). При возобновлении прежнего состояния или прежней внешней ситуации доминанта может снова возникнуть (актуализация доминанты). Например, в предстартовом состоянии активизируются все те нервные центры, которые входили в рабочую систему во время предыдущих тренировок, и, соответственно, усиливаются функции, связанные с работой. Мысленное выполнение физических упражнений или представление движений также воспроизводит рабочую доминанту, что обеспечивает тренирующий эффект представления движений ияв/гяется основой так называемой идеомоторной тренировки. При полном расслаблении (напр, при аутогенной тренировке) спортсмены добиваются устранения рабочих доминант, что ускоряет процессы восстановления.
Как фактор поведения, доминанта связана с высшей нервной деятельностью и психологией чсловека^Цоминантаявляется физиологической основой акта внимания. При наличии доминанты многие 'влияния внешней среды остаются вне нашего внимания, но зато бо- дёе интенсивно улавливаются и анализируются те, которые нас особенно интересуют. Таким образом, доминанта является мощным фактором отбора биологически и социально наиболее значимых раздражений. -
3.5. ФУНКЦИИ СПИННОГО МОЗГА И ПОДКОРКОВЫХ ОТДЕЛОВ ГОЛОВНОГО МОЗГА
В ЦНС различают более древние сегментарныеи эволюционно более молодые надсегментарные отделы нервной системы. К сегментарным отделам относят спинной, продолговатый и средний мозг, участки которых регулируют функции отдельных частей тела, лежащих на том же уровне. Надсегментарные отделы—промежуточный мозг, мозжечок и кора больших полушарий не имеют непосредственных связей с органами тела, а управляют их деятельностью через нижележащие сегментарные отделы.
3.5.1. СПИННОЙ МОЗГ
Спинной мозг является низшим и наиболее древним отделом ЦНС. В составе серого вещества спинного мозга человека насчитывают около 13.5 млн. нервных клеток. Из них основную массу (97%) представляют промежуточные клетки (вставочные или интернейроны), которые обеспечивают сложные процессы координации внутри спинного мозга. Среди мотонейронов спинного мозга выделяют крупные альфа-Мотонейроны имелкие —гамма-мото- нейрон ы. От альфа-мотонейронов отходят наиболее толстые и быстропроводящие волокна двигательных нервов, вызывающие сокращения скелетных мышечных волокон. Тонкие волокна гамма- мотонейронов Не вызывают сокращения мышц. Они подходят к про- приорецепторам — мышечным веретенам и регулируют их чувствительность.
Рефлексы спинного мозга можно подразделить на двигательные, осуществляемые альфа-мотонейронами передних рогов, и вегетативные, осуществляемые афферентными клетками боковых рогов.
Мотонейроны спинного мозга иннервируют все скелетные мышцы (за исключением мышцлица). Спинной мозг осуществляет элементарные двигательные рефлексы — сгибательные и разгибательные, ритмические, шагательные, возникающие при раздражении кожи или проприорецепторов мышц и сухожилий, а также посылает постоянную импул ьсацию к мышцам, поддерживая мышечный тонус. Специальные мотонейроны иннервируют дыхательную мускулатуру—межреберные мышцы и диафрагму, и обеспечивают Дыхательные движения. Вегетативные нейроны иннервируют все внутренние органы (сердце, сосуды, потовые железы, железы внутренней секреции, пищеварительный тракт, мочеполовую систему).
Проводниковая функция спинного мозга связана с передачей в вышележащие отделы нервной системы получаемого с периферии потока информации и с проведением импульсов, идущих из головного мозга в спинной.
За последние годы разработаны специальные методики для изучения деятельности спинного мозга у здорового человека. Так, например, функциональное состояние альфа-мотонейронов оценивают по изменению ответных потенциалов мышц при периферических раздражениях—так называемому Н-рефлексу (рефлексу Гофмана) икроножной мышцы при раздражении большеберцового нерва и по Т-рефлексу (от тендон — сухожилие) камбаловидной мышцы при раздражении ахиллова сухожилия. Разработаны методики регистрации (с неповрежденных покрововтела) потенциалов, проходящих по спинному мозгу в головной.
3.5.2. ПРОДОЛГОВАТЫЙ МОЗГ И ВАРОЛИЕВ МОСТ
Продолговатый мозги варолиев мост (в целом — задний мозг) являются частью ствола мозга. Здесь находится большая группа черепномозговых нервов (от V до XII пары), иннервирующих кожу, слизистые оболочки, мускулатуру головы и ряд внутренних органов (сердце, легкие, печень). Тут же находятся центры многих пищевари - тельных рефлексов—жевания, глотания, движений желудка и части кишечника, выделения пищеварительных соков, а также центры ие- которых защитных рефлексов (чихания, кашля, мигания, слезоотделения, рвоты) и центры водно-солевого и сахарного обмена, Надне IV желудочка в продолговатом мозге находится жизненно важный дыхательный центр, состоящий из центров вдоха и выдоха. Его состав* ляютмелкие клетки, посылающие импульсы кдыхательным мышцам через мотонейроны спинного мозга.
В непосредственной близости расположен сердечно-сосудистый центр. Его крупные клетки регулируют деятельность сердца и просвет сосудов. Переплетение клеток дыхательного и сердечно-сосудистого центров обеспечивает их тесное взаимодействие.
Продолговатый мозг играет важную роль в осуществлении двигательных актов и в регуляции тонуса скелетных мышц, повышая тонус мышц-разгибателей. Он принимает участие, в частности, в осуществ- лении установочных рефлексов позы (шейных, паЪщитн\лх), Через продолговатый мозг проходят восходящие пути слуховой, веспшбуляр- ной, проприоцептивной и тактильной чувствительности.
3.5.3. СРЕДНИЙ МОЗГ
В состав среднего мозга всходят четверохолмия, черная субстанция и красные ядра. Впередних буфах четверохолмия находятся зрительные подкорковые центры,авзааних— слуховые. Средний мозг
участвует в регуляции движений глаз, осуществляет зрачковыйрефлекс (расширение зрачков в темноте и сужение их на свету).
Четверохолмия выполняют ряд реакций, являющихся компонентами ориентировочного рефлекса. В ответ на внезапное раздражение происходит поворот головы и глаз в сторону раздражителя, а у животных — настораживание ушей. Этот рефлекс (по И. П. Павлову, • рефлекс «Что такое?») необходим для подготовки организма к своевременной реакции налюбое новое воздействие.
Черная субстанция среднего мозга имеет отношение к рефлексам жевания и глотания, участвует в регуляции тонуса мышц (особенно при выполнении мелких движений пальцами рук) и в организации содружественныхдвигательных реакций.
Красное ядро среднего мозга выполняет моторные функции — регулирует тонус скелетных мышц, вызывая усиление тонуса мышц- сгибателей. Оказывая значительное влияние на тонус скелетных мышц, средний мозгпринимаетучастие в рядеустановочныхрефлек- ' сов поддержания позы (выпрямительных — установке тела теменем вверх и др.).
3.5.4. ПРОМЕЖУТОЧНЫЙ МОЗГ
В состав промежуточного мозга входят таламус (зрительные бугры) и гипоталамус (подбугорье).
Через таламус проходят все афферентные пути (за исключе- нием обонятельных), которые направляются в соответствующие воспринимающие области коры (слуховые, зрительные и пр.). Ядра таламуса подразделяются на специфические и неспецифические. К специфическим относят переключательные (релейные) ядра и ассоциативные. Через переключательные ядра таламуса передаются афферентные влияния от всех рецепторов тела. Ассоциативные ядра получают импульсы от переключательных ядер и обеспечивают их взаимодействие. Помимо этих ядер в таламусе имеются неспецифические ядра, которые оказывают как активирующие, так и тормозящие влияния на небольшие области коры.
Благодаря обширным связям таламус играет важнейшую роль в жизнедеятельности организма. Импульсы, идущие от таламуса в кору, изменяют состояние корковых нейронов и регулируют ритм корковой активности. С непосредственным участием таламуса происходит образование условных рефлексов и выработка двигательных навыков, формирование эмоций человека, его мимики. Таламусу принадлежит большая роль в возникновении ощущений, в частности ощущения боли. С его деятельностью связыва- ютрегуляцию биоритмов в жизни человека (суточных, сезонных и др.).
Г ипоталамус является высшим подкорковым центрам регуляции вегетативных функций, состояний бодрствования и сна. Здесь расположены вегетативные центры, регулирующие обмен вещества организме, обеспечивающие поддержание постоянства температуры тела (у теплокровных) и нормального уровня кровяного давления, поддерживающие водный баланс, регулирующие чувство голода и насыщения. Раздражения задних ядер гипоталамуса вызывает усиление симпатических влияний, а передних — парасимпатические эффекты.
Благодаря связи гипоталамуса с гипофизом (гипоталамо-ги- пофизарная система) осуществляется контроль деятельности желез внутренней секреции. Вегетативные и гормональные реакции, регулируемые гипоталамусом, являются компонентами эмоциональных и двигательных реакций человека.
Дата добавления: 2015-05-21; просмотров: 826;