ВВЕДЕНИЕ. ИСТОРИЯ ФИЗИОЛОГИИ 3 страница
3.5.5. НЕСПЕЦИФИЧЕСКАЯ СИСТЕМА МОЗГА
Неспецифическая система занимает срединную часть ствола мозга. Она не связана с анализом какой-либо специфической чувствительности или с выполнением определенных рефлекторных реакций. Импульсы в эту систему поступают через боковые ответвления от всех специфических путей, в результате чего обеспечивается их обширное взаимодействие. Для неспецифической системы характерно расположение нейронов в виде диффузной сети, обилие и разнообразие их отростков. В связи с этим она и получила название сетевидного образования или ретикулярной формации.
Разл ичают два типа влияния неспецифической системы на работу других нервных центров — активирующее и тормозящее. Оба типа этих влияний могут быть восходящими (к вышележащим центрам) и нисходящими (к нижележащим центрам). Они служат для регулирования функционального состояния мозга, уровня бодрствования и регуляции позно-тонических и фазных реакций скелетных мышц.
3.5.6. МОЗЖЕЧОК
Мозжечок — это надсегментарное образование, не имеющее непосредственных связей с исполнительными аппаратами. Мозжечок состоит из непарного образования—червя и парных полушарий.
Основными нейронами коры мозжечка являются многочисленные клетки Пуркинье. Благодаря обширным связям (на каждой клетке оканчивается до 200 ООО синапсов) в них происходит интеграция самых различных сенсорных влияний, в первую очередь проприо- цептивных, тактильных и вестибулярных. Представительство разных периферических рецепторов в коре мозжечка имеет соматото- пическую организацию (греч. соматос—тело, топос—место), т. е. отражает порядок их расположения в теле человека. Кроме того, этот порядок расположен ия соответствует такому же порядку расположения представительства участков тела в коре больших полушарий, что облегчает обмен информацией между корой и мозжечком и обеспечивает их совместную деятельность в управлении поведением человека. Правильная геометрическая организация нейронов мозжечка обусловливает его значение в отсчете времени и четком поддержании темпа циклических движений.
Основной функцией мозжечка является регуляция позно-тони- ческихреакций и координация двигательной деятельности (Орбе- лиЛ. А., 1926).
По анатомическим особенностям (связям коры мозжечка с его ядрами) и функциональному значению мозжечок подразделяют на три продольные зоны:
• внутреннюю или медиальную —кору червя, функцией которой является регуляция тонуса скелетных мышц, поддержание позы и равновесия тела;
• промежуточную —среднюю часть коры полушарий мозжечка, функция которой состоит в согласовании позных реакций сдвижениями и коррекции ошибок;
• боковую или латеральную кору полушарий мозжечка, которая совместно с промежуточным мозгом и корой больших полушарий участвует в программировании быстрых баллистическихдвижений (бросков, ударов, прыжков и пр.).
3.5.7. БАЗАЛЬНЫЕ ЯДРА
Кбазальным ядрам относят полосатое тело, состоящее изхвоста- того ядра и скорлупы, и бледное ядро, а в настоящее время причисляют также миндалевидное тело (относящееся к вегетативным центрам лимбической системы) и черную субстанцию сред него мозга.
Афферентные влияния приходят к базал ьным ядрам от рецепторов тела через таламус и от всех областей коры бол ьших полушарий. Они почти исключительно поступают в полосатое тело. Эфферентные влияния от него направляются к бледному ядру и далее к стволовым центрам экстрапирамидной системы, атакже через таламус обратно к коре.
Базал ьные ядра участвуют в образовании условных рефлексов и осуществлении сложных безусловных рефлексов (оборонительных, пище- добывательных и др.). Они обеспечивают необходимое положение тела во время физической работы, атакже протекание автоматических ритмических движений (древних автоматизмов).
Бледное ядро выполняет основную моторную функцию, а полосатое тело регулирует его активность. В настоящее время выявлено значение хвостатого ядра в контроле сложных психических процессов—тптаыш, памяти, обнаружении ошибок.
3.6. ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА
Все функции организма условно можно разделить на соматические, или анимальные (животные), связанные с восприятием внешней информации и деятельностью мышц, и вегетативные (растительные), связанные с деятельностью внутренних органов, — процессы дыхания, кровообращения, пищеварения, выделения, обмена веществ, роста и размножения.
3.6.1. ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Вегетативной нервной системой называют совокупность эфферентных нервных клеток спинного и головного мозга, а также клеток особых узлов (ганглиев), иннервирующих внутренние органы. Раздражения различных рецепторов тела могут вызвать изменения как соматических, так и вегетативных функций, так как афферентные и центральные отделы этих рефлекторных дуг общие. Они различаются лишь своими эфферентными отделами. Характерной особенностью эфферентных путей, входящих в рефлекторные дуги вегетативныхрефлексов, является их двухнейронное строение (один нейрон находится в ЦНС, другой — в ганглиях или в иннервируемом органе).
Вегетативная нервная система подразделяется на два отдела — симпатический и парасимпатический (рис. 5).
Эфферентные пути симпатической нервной систе- м ы начинаются в грудном и поясничном отделах спинного мозга от нейронов его боковых рогов. Передача возбуждения с предузловых симпатических волокон на послеузловые происходит с участием медиатора ацетилхолина, а с послеузловых волокон на иннервируемые органы — с участием медиатора норадреналина. Исключением являются волокна, иннервирующие потовые железы и расширяющие сосуды скелетных мышц, где возбуждение передается с помощью ацетилхолина.
Эфферентные пути парасимпатической нервной с и ст е м'ъг начинаются втоловном мозге — от некоторых ядер среднего и продолговатого мозга, и в спинном мозге у— от нейронов крестцового отдела. Проведение возбуждения в синапсах парасимпатического пути происходит с участием медиатора ацетилхолина. Вто-
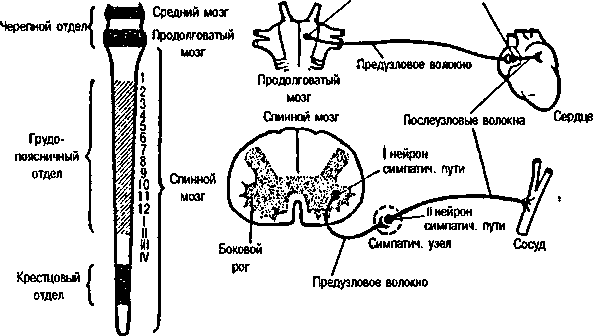 Рис. 5. Вегетативная нервная система
Слева — область выхода волокон; парасимпатической (черный цвет) и симпатической (заштриховано) систем.
Справа — строение эфферентной части рефлекторной дуги вегетативных
рефлексов. Слева — схема среднего, продолговатого и спинного мозга.
Арабские цифры — номера грудных сементов, римские — номера поясничных сегментов.
Рис. 5. Вегетативная нервная система
Слева — область выхода волокон; парасимпатической (черный цвет) и симпатической (заштриховано) систем.
Справа — строение эфферентной части рефлекторной дуги вегетативных
рефлексов. Слева — схема среднего, продолговатого и спинного мозга.
Арабские цифры — номера грудных сементов, римские — номера поясничных сегментов.
|
рой эфферентный нейрон находится в иннервируемом органе или вблизи от него.
Высшим регулятором вегетативных функций является гипоталамус, который действует совместно с ретикулярной формацией и лимбической системой под контролем коры больших полушарий. Кроме того, нейроны, расположенные в самих органах или в симпатических узлах, могут осуществлять собственные рефлекторные реакци и без участия ЦНС — «периферическиерефлексы».
3.6.2. ФУНКЦИИ СИМПАТИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
С участием симпатической нервной системы протекают многие важные рефлексы в организме, направленные на обеспечение его деятельного состояния, в том числе — его двигательной активности. К ним относятся рефлексырасширения бронхов, учащения иусиления сердечных сокращений, расширения сосудов сердца и легких при одновременном сужении сосудов кожи и органов брюшной полости (обеспечениеперераспределения крови), выброс депонированной крови из печени и селезенки, расщепление гликогена до глюкозы в печени (мобилизация углеводных источников энергии), усиление деятельности желез внутренней секреции и потовых желез. Симпатическая нервная система снижает деятельность ряда внутренних органов: и результате сужения сосудов в почках уменьшаются процессы моче- образования, угнетается секреторная и моторная деятельность органов желудочно-кишечного тракта; предотвращается актмочеиспус- кания—расслабляется мышца стенки мочевого пузыря и сокращается его сфинктер.
Повышенная активность организма сопровождается симпатическим рефлексом расширения зрачка. Огромное значение для двига- тел ьной деятельности организма и меет трофическое влияние симпатических нервов на скелетные мышцы, улучшающее их обмен веществ и функциональное состояние, снимающее утомление.
Симпатический отдел нервной системы не тол ько повышает уровень функционирования организма, но и мобилизует его скрытые функциональные резервы, активирует деятельность мозга, повышает защитные реакции (иммунные реакции, барьерные механизмы и др.), запускает гормональные реакции. Особенное значение имеет симпатическая нервная система при развитии стрессовых состояний, в наиболее сложныхусловияхжизнедеятельности. J1. А. Орбели подчеркивал важнейшее значение симпатических влияний для приспособления (адаптации) организма к напряженной работе, к различным условиям внешней среды. Эта функция была им названа адаптационно-трофической.
3.6.3. ФУНКЦИИ ПАРАСИМПАТИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
Парасимпатическая нервная система осуществляет сужение бронхов, замедление и ослабление сердечных сокращений; сужение сосудов сердца; пополнение энергоресурсов (синтез гликогена в печени и усиление процессов пищеварения); усиление процессов моче образования в почках и обеспечение акта мочеиспускания (coKpamei I ие мышц мочевого пузыря и расслабление его сфинктера) и др. Парасимпатическая нервная система преимущественно оказывает пусковые влияния: сужение зрачка, бронхов, включение деятельности пищеварительных желез и т. п.
Деятельность парасимпатического отдела вегетативной нервной системы направлена на текущую регуляцию функционального состояния, на поддержание постоянства внутренней среды — гомеостаза. Парасимпатический отдел обеспечивает восстановление различных физиологических показателей, резко измененных после напряженной мышечной работы, пополнение израсходованных энергоресурсов. Медиатор парасимпатической системы — ацетил- холин, снижая чувствительность адренорецепторов к действию адреналина и норадреналина, оказывает определенное анти- стрессорное влияние.
Через вегетативные симпатические и парасимпатические пути ЦНС осуществляет некоторые вегетативные рефлексы, начинающиеся с различных рецепторов внешней и внутренней среды: висцеро-висцеральные (с внутренних органов на внутренние органы — например, дыхательно-сердечный рефлекс); дер- мо-висцеральные (с кожных покровов — изменение деятельности внутренних органов при раздражении активных точек кожи, например, иглоукалыванием, точечным массажем); с рецепторов глазного яблока — глазо-сердеч- ный рефлекс Ашнера(уреже- ние сердцебиений при надавливании на глазные яблоки — парасимпатический эффект); моторно-висцеральные — например, о р- то статичес к а я проба (учащение сердцебиения при переходе из положения лежа в положение стоя — симпатический эффект) и др. (рис. 6). Они используются для оценки функционального состояния организма и особенно состояния вегетативной нервной системы (оценки влияния симпатического или парасимпатического ее отдела).
| ! л v ... I |
| 3.6.4. ВЕГЕТАТИВНЫЕ РЕФЛЕКСЫ |
| Рис. 6. Вегетативные рефлексы Влияние положения тела на частоту сердечных сокращений (уд./мин). (По: Могендович М.Р., 1972) |
3.7. ЛИМБИЧЕСКАЯ СИСТЕМА
Под лимбической системой понимают ряд корковых и подкорковых структур, функции которых связаны с организацией мотивационно-эмоциональных реакций, процессами памяти и обучения.
Корковые отделы лимбической системы, представляющие ее высший отдел, находятся на нижних и внутренних поверхностях больших полушарий (участки лобной коры, поясная извилина или лимбическая кора, гиппокамп идр.). К подкорковым структурам лимбической системы относят гипоталамус, некоторые ядра таламуса, среднего мозга и ретикулярной формации. Между всеми этими
отделами имеются тесные прямые и обратные связи, образующие так называемое «лимбическое кольцо».
Лимбическая система участвует в самых разнообразных nponeie- ниях деятельности организма—в регуляции пищевого и питьевого поведения, цикла сон-бодрствование, в процессах формирования памятного следа (отложения и извлечения из памяти), в развитии агрессивно-оборонительных реакций, обеспечивая избирательный характер поведения. Она формирует положительные и отрицательные эмо- циисо всеми двигательными, вегетативными и гормональными их компонентами. Электрические раздражения различных участков лимбической системы через вживленные электроды (в экспериментах на животных, в клинике при лечении больных) выявили наличие центров удовольствия, формирующих положительные эмоции, и неудовольствия, формирующих отрицательные эмоции. Изолированное раздражение таких точек в глубоких структурах мозга человека вызывало появление чувства «беспричинной радости», «беспредметной тоски», «безотчетного страха».
3.8. ФУНКЦИИ КОРЫ БОЛЬШИХ ПОЛУШАРИЙ
У высших млекопитающих животных и человека ведущим отделом ЦНС является кора больших полушарий.
3.8.1. КОРКОВЫЕ НЕЙРОНЫ
Кора представляет собой слой серого вещества толщиной 2-3 мм, содержащий в среднем около 14 млрд. нервных клеток. Характерным в ней является обилие межнейронных связей, рост которых продолжается до 18 лет, а в ряде случаев и далее.
Основными типами корковых клеток являются пирамидные и звездчатые нейроны. Звездчатые нейроны связаны с процессами восприятия раздражений и объединением деятельности различных пирамидных нейронов.
Пирамидные нейроны осуществляют эфферентную функцию коры (преимущественно через пирамидный тракт) и внут- рикорковые процессы взаимодействия между удаленными друг от друга нейронами. Наиболее крупные пирамидные клетки — гигантские пирамиды Беца находятся в передней центральной извилине (моторной зоне коры).
Функциональной единицей коры является вертикальная колонка взаимосвязанныхнейронов. Вытянутые по вертикали крупные пирамидные клетки с расположенными над ними и под ними нейронами образуют функциональные объединения нейронов. Все нейроны вертикальной колонки отвечают на одно и то же афферент
ное раздражение (от одного и того же рецептора) одинаковой реакцией и совместно формируют эфферентные ответы пирамидных нейронов. По мере надобности вертикальные колонки могут объединяться в более крупные образования, обеспечивая сложные реакции.
3.8.2. ФУНКЦИОНАЛЬНОЕ ЗНАЧЕНИЕ РАЗЛИЧНЫХ КОРКОВЫХ ПОЛЕЙ
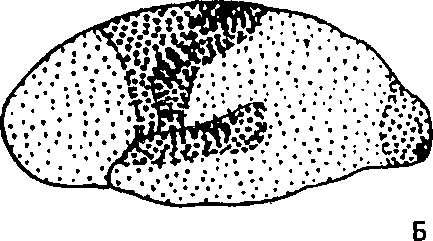
По особенностям строения и функциональному значению отдельных корковых участков вся кора подразделяется на три основные группы полей—первичные, вторичные и третичные (рис.7).
Первичные поля связаны с органами чувств и органами движения на периферии. Они обеспечивают возникновение ощущений. К ним относятся, например, поле болевой и мышечно-суставной чувствительности в задней центральной извилине коры, зритель-
|
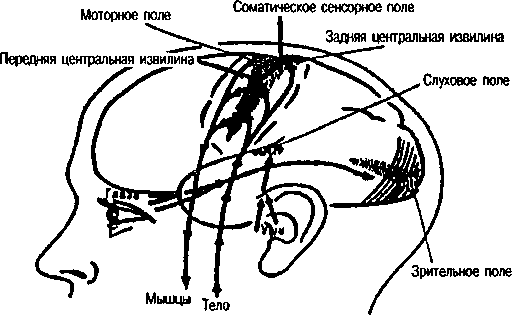 Рис. 7. Первичные, вторичные и третичные поля коры больших полушарий
На А: крупные точки — первичные поля, средние — вторичные поля, мелкие — третичные поля (по Г.И. Поляков 1964, А.Р. Лурия, 1971)
На Б: первичные (проекционные) поля коры больших полушарий (по В.Пенфильд, Л. Робертс, 1964)
Рис. 7. Первичные, вторичные и третичные поля коры больших полушарий
На А: крупные точки — первичные поля, средние — вторичные поля, мелкие — третичные поля (по Г.И. Поляков 1964, А.Р. Лурия, 1971)
На Б: первичные (проекционные) поля коры больших полушарий (по В.Пенфильд, Л. Робертс, 1964)
|
А
ное поле в затылочной области, слуховое поле в височной области и моторное поле в передней центральной извилине. В первичных полях находятся высокоспециализированные клетки-определители или детекторы, избирательно реагирующие только на определенные раздражения. Например, в зрительной коре имеются нейроны-детекторы, возбуждающиеся только при включении или при выключении света, чувствительные лишь к определенной его интенсивности, к конкретным интервалам светового воздействия, копределенной длине волны и т.д. При разрушении первичных полей коры возникают так называемые корковая слепота, корковая глухота а пип.
Вторичные поля расположены рядом с первичными. В них происходит осмысливание и узнавание звуковых, световых и других сигналов, возникают сложные формы обобщенного восприятия. При поражении вторичных полей сохраняется способность видеть предметы, слышать звуки, но человек их не узнает, не помнит значения.
Третичные поля развиты практически только у человека. Это ассоциативные области коры, обеспечивающие высшие формы анализа и синтеза и формирующие целенаправленную поведенческую деятельность человека. Третичные поля находятся в задней половине коры—междутеменными, затылочными и височными областями, и в передней половине — в передних частях лобных областей. Их роль особенно велика в организации согласованной работы обоих полушарий. Третичные поля созревают у человека позже других корковых полей и раньше другихдеградируют при старении.
Функцией задних третичных полей (главным образом, нижнетеменных областей коры) является прием, переработка и хранение информации. Они формируют представление о схеме тела и схеме пространства, обеспечивая пространственную ориентацию движений. Передние третичные поля (передне-лобные области) выполняют общую регуляцию сложных форм поведения человека, формируя намерения и планы, программы произвольных движений и контроль за их выполнением. Развитие третичных полей у человека связывают с функцией речи. Мышление (внутренняя речь) возможно только при совместной деятельности различных сенсорных систем, объединение информации от которых происходит в третичных полях. При врожденном недоразвитии третичных полей человек не в состоянии овладеть речью (произноситлишьбессмыслен- ные звуки) и даже простейшими двигательными навыками (не может одеваться, пользоваться орудиями труда и т. п.).
3.8.3. ПАРНАЯ ДЕЯТЕЛЬНОСТЬ И ДОМИНИРОВАНИЕ ПОЛУШАРИЙ
Обработка информации осуществляется в результате парной деятельности обоих полушарий головного мозга. Однако, как правило, одно из полушарий является ведущим — доминантным. У большинства людей с ведущей правой рукой (правшей) доминантным является левое полушарие, а соподчиненным (субдоминантным) — правое полушарие.
Левое полушарие по сравнению с правым имеет более тонкое нейронное строение, большее богатство взаимосвязей нейронов, более концентрированное представительство функций и лучшие условия кровоснабжения. В левом доминантном полушарии находится моторный центр речи (центр Брока), обеспечивающий речевую деятельность, и сенсорный центр речи, осуществляющий понимание слов. Левое полушарие специализировано на тонком сенсомоторном контроле за движениями рук.
У человека различаюттри формы функциональной асим- м е т р и и: моторную, сенсорную и психическую. Как правило, у человека имеются ведущая рука, нога, глаз и ухо. Однако проблема функциональной асимметрии довольно сложна. Например, у человека-правши может быть ведущим левый глаз или левое ухо, сигналы от которых являются главенствующими. При этом в каждом полушарии могут быть представлены функции не только противоположной, но и одноименной сторонытела. В результате этого обеспечивается возможность замещения одного полушария другим в случае его повреждения, а также создается структурная основа для переменного доминирования полушарий при управлении движениями.
Психическая асимметрия проявляется в виде определенной специализации полушарий. Для левого полушария характерны аналитические процессы, последовательная обработка информации, в том числе с помощью речи, абстрактное мышление, оценка временных отношений, предвосхищение будущих событий, успешное решение вербально-логических задач. В правом полушарии информация обрабатывается целостно, синтетически (без расчленения на детали), с учетом прошлого опыта и без участия речи, преобладает предметное мышление. Эти особенности позволяют связывать с правым полушарием восприятие пространственных признаков и решение зрительно-пространственных задач. Функции правого полушария связаны с прошедшим временем, а левого — с будущим.
3.8.4. ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ КОРЫ БОЛЬШИХ ПОЛУШАРИЙ
Изменения функционального состояния коры отражаются в записи ее электрической активности — электроэнцефалограммы (Э Э Г). Современные элекгроэнцефалографы усиливают
потенциалы мозга в 2-3 млн. раз и дают возможность исследовать ЭЭГ от многих точек коры одновременно, т. е. изучать системные процессы. Регистрация ЭЭГ производится в виде чернильной записи на бумаге, а также в виде целостной картины на схеме поверхности мозга, т. е. карты мозга (метод картирования) на экране мониторов современных компьютерных систем (рис. 8).
Различают определенные диапазоны частот, называемые ритмами ЭЭГ (рис. 9): в состоянии относительного покоя чаше всего регистрируется альфа-ритм (8-13 колебаний в 1 с); в состоянии активного внимания — бета-ритм (14 колебаний в 1 с и выше); при за-
 Рис. 8, Картирование мозга Многоканальная регистрация электроэнцефалограммы (ЭЭГ) человека на экране монитора и отражение возбужденных (светные зоны) и заторможенных (темные зоны) участков коры
Рис. 8, Картирование мозга Многоканальная регистрация электроэнцефалограммы (ЭЭГ) человека на экране монитора и отражение возбужденных (светные зоны) и заторможенных (темные зоны) участков коры
|
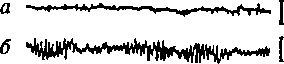
|
| I |
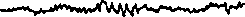
|
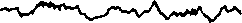
|
| 50мкв |
| I |
в
г
•| q эимкв
е VVVMMM*»I ж \zVyVVVV44f\zvVv/' I
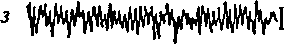
|
Рис. 9. ЭЭГзатылочной (а—д) и моторной (е—з) областей коры больших полушарий человека при различных состояниях и во время мышечной работы а — активное состояние, глаза открыты (бета-ритм); б— покой, глаза закрыты (альфа-ритм); в — дремота (тета-ритм г — засыпание; д — глубокий сон (дельта-ритм); е — непривычная или тяжелая работа — асинхронная частая активность (явление десинхронизации); ж — циклическая работа — медленные потенциалы в темпе движений («меченые ритмы» ЭЭГ); з — выполнение освоенного движения — появление альфа-ритма.
сыпании, некоторых эмоциональных состояниях—тета-ритм (4-7 колебаний в 1 с); при глубоком сне, потере сознания, наркозе — дельта-ритм (1-3 колебания в 1 с).
В ЭЭГотражаются особенности взаимодействия корковых нейронов при умственной и физической работе (Ливанов М. Н., 1972). Отсутствие налаженной координации при выполнении непривычной или тяжелой работы приводит ктак называемой десинхронизации ЭЭГ — быстрой асинхронной активности. По мере формирования двигательного навыка в ЭЭГ возникают явления синхронизации ЭЭГ—усиления взаимосвязанности (синхронности и синфазности) электрической активности различных областей коры, участвующих вуправлении движениями. При циклической работе появляются медленные потенциалы в темпе выполняемого, воображаемого или предстоящего движения — «меченые р и т- м ы» (Сологуб Е. Б., 1973).
П ом им о фоновой активности в ЭЭ Г выделяют отдельные потенциалы, связанные с какими-либо событиями: вызванные потенциалы, возни кающие в ответ на внеш ние раздражения (слуховые, зрительные и др.); потенциалы, отражающие мозговые процессы при подготовке, осуществлении и окончании отдельных двигательных актов — это «волна ожидания» или условная негативная волна (Уолтер Г., 1966), премоторные, моторные и финальные потенциалы и др. Кроме того, регистрируют различные сверхмедленные колебания: длительностью от нескольких секунд до десятков минут (в частности, так называемые «омега-по- тенциалы» и др.), которые отражаютбиохимические процессы регуляции функций и психической деятельности,
4. ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ
Развивая идеи И. М. Сеченова о рефлекторной основе поведен- ческойдеятельности целостного организма, И. П. Павлов пришел к мысли, что в изменяющихся условиях внешней среды недостаточно обладать стандартными рефлекторными реакциями, атребуется выработка новых рефлексов, адекватных новым условиям существования. Впервые об условных рефлексах он заговорил в известной Мадридской речи в 1903г.
4.1. УСЛОВИЯ ОБРАЗОВАНИЯ И РАЗНОВИДНОСТИ УСЛОВНЫХ РЕФЛЕКСОВ
Условные рефлексы по многим признакам отличаются от безусловных (табл. 1).
Таблица I Различия условных и безусловных рефлексов
Безусловные рефлексы
Врожденные реакции Постоянно существующие реакции
Видовые рефлексы
Имеются готовые рефлекторные
дуги
Осуществляются всеми отделами ЦНС
Условные рефлексы
Приобретенные реакции Временно существующие реакции
Индивидуальные рефлексы Образуются новые рефлекторные дуги
Осуществляются ведущими отделами ЦНС
ровании новой рефлекторной дуги, в которой к эфферентной части бе- зусловного рефлекса присоединялась новое афферентное начало рефлекторной дуги, идущее от зрительных путей. Между центрами этих исходных рефлексов сформировалась новая связь, которую И. П. Павлов назвал рременной связью, так как в случае прекращения подачи пищи после светового сигнала слюнной условный рефлекс исчезал. В ходе выработки условного рефлекса наблюдались определенные фазы этого процесса:
1) генерализации (обобщенное восприятие сигнала, когда условная реакция наблюдалась на любой сходный сигнал), основой чего были процессы иррадиации возбуждения в коре больших полушарий;
2) концентрации возбуждения (реакция только на конкретный сигнал), что появлял осьза счет вырабатываемого условного торможения на посторонние неподкрепляемые сигналы и ^стабилизации (упрочения условного рефлекса).
В дальнейших исследованиях условные рефлексы были выработаны в разнообразных экспериментальных условиях (в том числе в условиях свободного поведения) у различных животных, птиц, рыб, черепах, даже у амеб. Изучение биопотенциалов коры больших полушарий показало, что условием образования временной связи между изучаемыми корковыми центрами является пространственная синхронизация их электрической активности.
Различают несколько разновидностей условных рефлексов: 1) натуральные —на сигналы, характеризующие безусловные раздражители (напр., запах мяса для слюнного рефлекса) и искусственные— на посторонние сигналы (например, запах мяты); 2) наличные и следовые на условный сигнал, непосредственно предшествующий безусловному подкреплению, и на его следовое влияние; 3)положительные (с активным проявлением ответной реакции) и отрицательные (с ее торможением); 4) условные рефлексы на время —> при ритмической подаче условных сигналов ответная реакция появляется через заданный интервал даже при отсутствии очередного сигнала; 5) условные рефлексы первого порядка — на один предшествующий условный раздражитель — и более высоких порядков, когда безусловному подкреплению предшествует сочетание двух последовательно подающихся сигналов (свет + звук) — условный рефлекс второго порядка, трех сигналов (свет+звук + касалка)—условный рефлекс третьего порядка и т. д. У собак вырабатываются, в основном, рефлексы третьего порядка, у обезьян — четвертого, у грудного ребенка — 5-6 порядка, у взрослого человека — двадцатого и более порядков. Освоение речи человеком представляет собой формирование огромной цепи условно-безусловных рефлексов, не требующих специального подкрепления.
При формировании новых двигательных навыков возникают особые рефлексы, которые в отличие от сенсорных рефлексов или рефлексов Ipoda (в которых новой частью рефлекторной дуги была афферентная часть) имеют новую часть рефлекторной дуги в эфферентном отделе (новые исполнительные аппараты — мышцы). Это так называемые инструментальные или оперантныерефлексы — рефлексы II рода (Ко] юрский Ю. М., 1970).
4.2. ВНЕШНЕЕ И ВНУТРЕННЕЕ ТОРМОЖЕНИЕ УСЛОВНЫХ РЕФЛЕКСОВ
По своему происхождению торможение условных рефлексов может быть безусловным (врожденным) и условным (выработанным в течение жизни). К безусловному торможению относят охранительное или запредельное торможение, возникающее при чрезмерно сильном или длительном раздражении, и внешнеетормо- жение условных рефлексов посторонними для центров условного рефлекса раздражителями (например, нарушение непрочного двигательного навыка у спортсмена в необычных условиях соревнований).
Условное торможение вырабатывается при отсутствии подкрепления условного сигнала. Различают несколько видов условного торможения: угасательное, дифференцировочное и запаздывающее. Угасание развивается при повторении условного сигнала без подкрепления. Например, имея прочный слюнный условный рефлекс у собаки на вспышку света и затем применяя свет без подкрепления, можно получить последовательно следующие условные ответы — 10,8,6,4,5,2,0,0,0 капель слюны.
Дата добавления: 2015-05-21; просмотров: 1160;