Основные данные о строении артериальной сети мягкой мозговой оболочки у кролика 2 страница
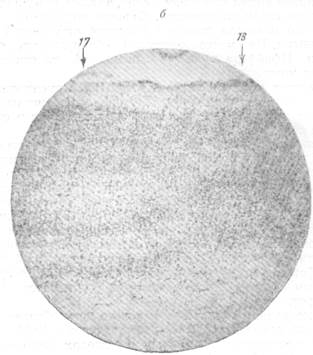
Соответствие ангиоархитектонического строения цитоархитектоническому
в коре головного мозга мартышки.
а— ангиоархитектоника полей 17 и 18 мозговой коры (инъекция тушью); б— цитоархи-тектоника тех же полей (окраска по Нисслю).
Нельзя, конечно, не согласиться с Пфайфером в том, что клеточное строение того или другого поля мозговой коры находит свое отражение в характере сосудисто-капиллярной сети данного участка. На препаратах с импрегнированной сосудистой сетью, например, можно вполне отчетливо различить два рядом расположенных поля, обладающих различным клеточным строением. На рис. 65 представлены поля 17 и 18 мозговой коры обезьяны, каждое из которых имеет характерное для него ангиоархитектоническое строение (рис. 65, а) соответственно размещению в нем клеточных элементов (рис. 65, б). Можно считать, что расположение клеточных слоев должно в известной мере определять способ отхождения боковых ветвей, образующих со стволом артерии различный угол в зависимости от направления клеточного слоя.
Однако справедливость основной мысли Пфайфсра о соответствии цитоархитектоники с ангиоархитектоникой в значительной мере подвергается сомнению в иллюстративной части его работы.
Так, на многочисленных фотографиях, приведенных с целью подтвердить теоретические положения автора, в качестве артерий повсюду фигурируют вены. Поэтому кровообращение описанных этим автором клеточных полей приходится расценивать не как артериальное снабжение их, а как венозный отток с этих участков.
Если же просмотреть артериальные стволы, зачастую располагающиеся на тех же микрофотографиях Пфайфера, то далеко не всегда можно установить отхождение боковых ветвей от основного ствола соответственно расположению слоев клеточного поля. В большинстве случаев, ветви образуют со стволом острый угол.
Это обстоятельство, по нашему мнению, должно указывать на то, что особенности цитоархитектоники того или иного поля коры взрослого человека или животных постоянно будут находить свое отражение в плотности и форме петель капиллярной сети, тогда как угол, под которым ветвь отходит от артериального ствола, не может быть характерным и постоянным.
Несомненно, решение вопроса о факторах, определяющих характер взаимного расположения клеточного и сосудистого компонентов в различных полях коры, нужно искать при изучении ранних стадий онтогенетического развития.
Работы, ведущиеся в этом направлении в нашей лаборатории, дали уже определенные результаты. Проследив формирование сосудисто-капиллярной сети в мозгу эмбрионов на различных стадиях организации клеточного строения, 3. Н. Киселева установила, что на ранней стадии эмбрионального развития человека или животных врастающие в мозговое вещество сосуды идут в радиальном направлении от поверхности переднего мозгового пузыря к полости его. При своем следовании через стенку пузыря сосуды располагаются параллельно друг другу и соединяются между собой поперечно идущими сосудами.
Благодаря описанному ходу и соединению сосудов в стенке мозгового пузыря организуется сосудистая есть своеобразного строения. Петли этой сети вытянуты в длину по ходу радиального сосуда. На импрегниро-ванных препаратах, приготовленных из эмбрионов ранних стадий развития, сосудистая сеть имеет вид сложной лестничной системы, перекладины в которой соединяют не только сосуды, располагающиеся в одной плоскости, но и сосуды, лежащие в различных вертикальных плоскостях (рис. 66).
Совершенно иная картина сосудистой сети наблюдается в матриксе. Прослеживая ее организацию, можно видеть, что она сформирована из
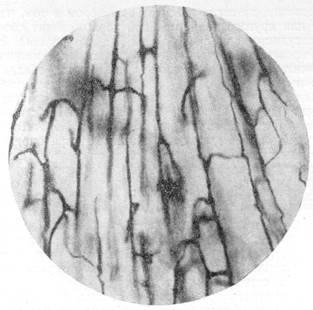
Рис. 66. Строение сосудистой сети в стенке переднего мозгового пузыря у эмбриона
кролика. Сосудистая сеть импрегнирована серебром по методу В. Н. Клосовского. Увеличение 100.
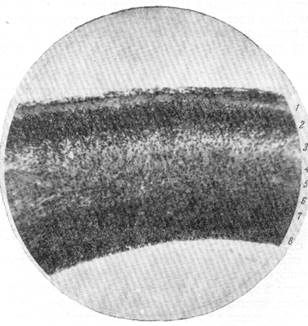
Рис. 67. Клеточное строение стенки переднего мозгового пузыря у эмбриона собаки.
Стадия 8 слоев.
1 — краевой покров; 2 — кора; 3 — промежуточный слой; 4 — наружный переходной слой; 5 — наружный полосатый слой; 6 — внутренний переходной слой; 7 — внутренний полосатый слой; 8 — матрикс. Окраска по методу Ниссля. Увеличение 22.
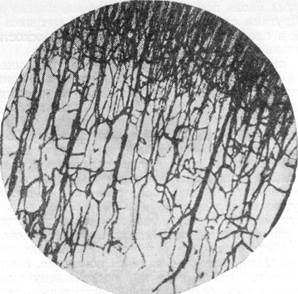
Рис. 68. Ветвление радиальной артерии на уровне
наружного переходного слоя у эмбриона собаки
длиной 11 см.
Обращает на себя внимание острый угол между стволом артерии и ее боковой ветвью. Импрегнация серебром. Увеличение 22.
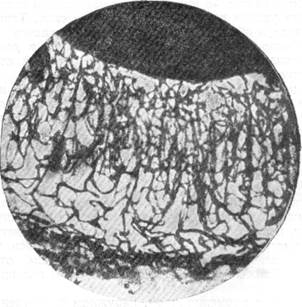
Рис. 69. Сосудисто-капиллярная сеть стенки переднего мозгового пузыря у эмбриона собаки длиной
11 см.
Стадия 8 слоев.
Импрегнация сосудистой сети серебром по методу Б. Н. Клосовского. Увеличение 22.
петель, вытянутых вдоль поверхности желудочка, плотно лежащих друг возле, друга. Другими словами, сосудистая сеть матрикса оказывается расположенной в плоскости, перпендикулярной к плоскости размещения сосудов в остальной части стенки мозгового пузыря.
На стадии эмбрионального развития, характеризующейся наличием четырехслойной стенки переднего мозгового пузыря, общий вид сосудистой сети значительно меняется. Так же как и на предыдущих этапах онтогенеза, радиальные сосуды проходят, не ветвясь, краевой покров, но в появившемся теперь корковом слое от них отходят боковые ветви, после дальнейших делений распадающиеся на капиллярную сеть.
Капиллярная сеть формирующейся коры состоит из петель вытянутой формы, длинная ось которых направлена по ходу радиальных сосудов. Сравнительно редкая капиллярная сеть с широкими петлями представляется все же более плотной при сравнении ее с соответствующей сетью промежуточного слоя.
В промежуточном слое сосудистая сеть сохраняет строение, описанное для более ранних стадий развития, т, е. состоит из очень широких и редких петель, образующихся в результате соединения радиально идущих сосудов с сосудами, направляющимися поперечно. Матрикс на стадии четырехслойной стенки мозгового пузыря попрежнему выделяется своей густой сосудистой сетью, расположенной параллельно поверхности желудочка.
При дальнейшем преобразовании стенки переднего мозгового пузыря и распределении клеточных масс на 8 слоев (рис. 67) не только увеличивается количество боковых ветвей и плотность капиллярной сети в корковом слое, но и появляются новые ветви, отходящие от основного ствола в более глубоких слоях формирующейся стенки.
Так, например, из рис. 68 видно, как от радиальной артерии отходит боковая ветвь на уровне наружного переходного слоя. Такие же ветви от артерий отходят в области расположения внутреннего переходного слоя. Соответственно указанному отхождению ветвей в данных слоях оформляются капиллярные сети, расположение петель в которых чрезвычайно напоминает капиллярную сеть матрикса.
На рис. 69 и 70 видно, что одновременно с появлением капиллярных сетей на новых уровнях стенки переднего мозгового пузыря произошло преобразование сетей в ранее существовавших слоях. Капиллярная сеть коры заняла значительно большее пространство, стала значительно плотнее. Зато капиллярная сеть в матриксе стала реже и расположена на меньшем пространстве. Капиллярная сеть в промежуточном слое также стала плотнее.
Поскольку на разбираемой стадии развития отчетливо выражена слоистость стенки переднего мозгового пузыря и соответствующее ей распределение капиллярных сетей, на данном этапе развития легче, чем на более ранних, проследить способ отхождения боковых ветвей от артерий и вен. Просматривая серии препаратов, мы убедились, что боковые ветви от артерий неизменно отходят под острым углом от основного ствола, тогда как боковые ветви вен оставляют ствол под прямым углом (рис. 68, 71).
Таким образом, в промежуточном, наружном и внутреннем полосатых слоях радиальные сосуды на стадии восьми слоев не дают разветвлений, тогда как корковый, наружный и внутренний переходные слои, а также матрикс содержат, в зависимости от слоя, большее или меньшее количество боковых ветвей и различной густоты капиллярные сети (рис. 69, 70). Ветвление радиальных сосудов, а также формирование ка-
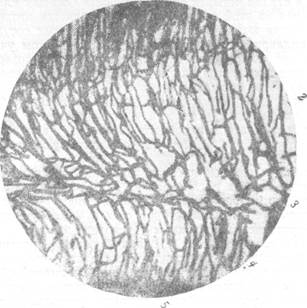
Рис. 70. Сосудисто-капиллярная сеть стенки мозгового пузыря на той же стадии
развития, что и на рис. 69.
1— кора; 2 — промежуточный слой; 3— наружный переходной, наружный полосатый и внутренний переходной слой: 4—внутренний полосатый слой; 5—матрикс. Увеличение 100.
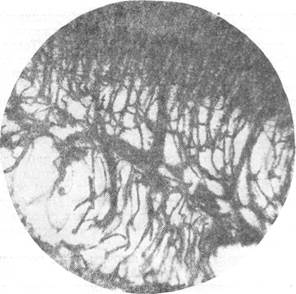
Рис. 71. Способ ветвления радиальной вены на уровне наружного переходного слоя
у эмбриона собаки длиной 11 см. Импрегнация серебром по методу Клосовского. Увеличение 22.
пиллярныхсетей находятся в прямом соответствии с расположением клеточных элементов на данном этапе развития.
Вернемся к стадии четырех слоев. Наиболее богатым клетками слоем стенки полушария на этой стадии онтогенеза является матрикс. Соответственно наибольшее количество ветвей от радиальных сосудов и наиболее густая капиллярная сеть отмечаются именно в этом участке станки полушария. То же можно сказать и о корковом слое, содержащем значительно меньшее количество клеточных элементов и менее густую с более широкими петлями капиллярную сеть.
Следующая стадия формирования стенки полушария (8 слоев) характеризуется увеличением мощности коркового слоя и возникновением внутреннего и наружного переходного слоя, где, наряду с матриксом, теперь сосредоточиваются большие массы мигрирующих и диференци-рующих клеток.
Одновременно с изменением распределения клеточного компонента произошло и изменение в распределении и степени организации сосудисто-капиллярной сети. Параллельно с увеличением мощности коркового слоя увеличилась густота капиллярной сети в нем. Возникли капиллярные сети во внутреннем и наружном переходном слое.
Таким образом, изменения в сосудистой системе протекают одновременно с преобразованиями клеточной организации стенки полушария.
Радиальные артерии прорастают из мягкой мозговой оболочки через всю толщу стенки полушария вплоть до матрикса и дают свои первичные разветвления в условиях беспорядочного расположения клеточных элементов в тех слоях стенки, через которые они проходят.
Следовательно, даже с точки зрения Пфайфера, угол отхождения боковых ветвей должен определяться здесь только мощностью боковой ветви и способ ветвления артерии на этих стадиях развития должен быть подобен способу, характерному для артерий в бесструктурной коре, например, в передней или задней центральной извилине человека и обезьяны.
Вполне вероятно, что при последующей диференцировке коры на слои преобразования сосудистого дерева будут выражаться в отхождении боковых ветвей соответственно расположению слоев. Тогда при наличии коры с параллельными слоями ветви артерий приобретут горизонтальное направление, являющееся, по мнению Пфайфера, особенностью ветвления артерий таких участков мозга, как gyrus fornicalus.
Но даже в таких случаях артерия, наряду с вторичными ветвями, отходящими от ствола под прямым углом, будет иметь первичные ветви, образующие с основным стволом острые углы, благодаря чему способ отхождения боковых ветвей будет весьма разнообразен.
Подобного рода артерии действительно можно наблюдать на препаратах. Одна, две, а то и несколько ветвей могут отходить от ствола под прямым углом, но большинство все же образует с ним острый угол.
Таким образом, надо признать, что попытка Пфайфера найти зависимость способа отхождепия ветвей артериальных стволов от характера расположения клеточных слоев не нашла себе подтверждения ни в онтогенезе, ни у взрослого животного и человека.
Все сказанное выше позволяет заключить, что, поскольку организация клеточного строения стенки полушария и сосудистой системы происходит одновременно, безусловно должно иметь место влияние одной системы на другую. На настоящем этапе развития науки можно лишь установить определенную зависимость степени развития капиллярной сети того или другого слоя какого-либо поля мозговой коры от количе-
ства находящихся в этом слое нервных клеток или синапсов [см. De Но, 1927; Пфайфер, 1940; Линдгрин (Lindgreen), 1940; 3. Н. Киселева, 1947]. Соотношение же расположения клеточных слоев и характера от-хождения боковых ветвей от снабжающих данное поле артерий требует дальнейших тщательных исследований.
Выяснение вопроса об ангиоархитектоническом строении полушарий головного мозга представляет сложную задачу. Решение ее требует изучения многих серий гистологических препаратов с представленной полностью сосудисто-капиллярной сетью, а также массы сопоставлений и сравнительных исследований. В литературе настоящего времени существуют пока всего две работы, авторы которых при решении одной и той же задачи пришли к противоположным выводам. Мы имеем в виду монографии Линдгрина (1940) и Пфайфера (1940).
Линдгрин (1940), работая методом Пиквортса на мозгах взрослых людей, пришел к заключению, что в большей части изокортекса ангиоар-хитектоника относительно однообразна и существуют лишь незначительные местные модификации. В отдельных полях коры, например, в височной доле, имеет место соответствие между ангиоархитектонической структурой и цито- и миэлоархитектоникой, в других же полях таких соотношений не наблюдается. Согласно данным этого исследователя, в мозгу можно различать поля, ангиоархитектоническая диференцировка которых выше диференцировки их в цито- и миэлоархитектонических отношениях, и наряду с этим поля, для которых эти соотношения обратны.
Одновременно с работой Линдгрина было опубликовано исследование Пфайфера (1940), изучавшего ангиоархитектоническое строение полушарий головного мозга обезьяны.
Используя метод инъекции сосудистого русла мозга раствором туши, Пфайфер пришел к выводу, что деление коры на основании ангиоархи-тектоничеокого строения ее может быть проделано со значительно большей точностью, чем деление коры соответственно расположению клеточных слоев в ней.
Наряду с этим в мозгу существуют также, области, которые по распределению в них сосудистой сети чрезвычайно трудно отграничить от рядом расположенных областей. На основе ангиоархитектонических картин коры головного мозга вполне возможно составление ангиоархи-тектонической карты, подобно картам, разработанным для нервных клеток и нервных волокон.
Рассматривая карты распределения сосудисто-капиллярной сети в коре полушарий головного мозга обезьяны, можно видеть, что предложенное Пфайфером деление коры на поля по ангиоархитектоническому признаку в основных чертах совпадает с цитоархитектоническими полями Экономо и Коскинаса. Однако в пределах цитоархитектонических полей нередко можно выделить добавочные более мелкие поля, отличающиеся по строению сосудистой сети от остальных областей того же поля, цито-архитектоника которого едина на всем его протяжении.
Ангиоархитектоника мозга, таким образом, является вопросом, разработка которого еще только начата. Потребуется немало исследований для того, чтобы представить строение сосудистой сети в различных отделах мозга хотя бы даже в общем виде.
В порядке последовательности изложения рассмотрим теперь существующие в настоящее время фактические данные относительно дальнейшего хода артерий и вен в сером и белом веществе.
Дюре, учение которого об анатомических основах мозгового кровообращения получило в свое время столь широкое распространение, впер-
вые в истории этого вопроса предложил деление артерий и вен мозга на короткие и длинные.
Короткие, или «корковые», по терминологии Дюре, артерии, по которым кровь поступает исключительно в серое вещество, дают при своем следовании через кору многочисленные ветви, делящиеся в свою очередь неоднократно вплоть до образования капилляров.
Капилляры формируют в сером веществе две сети. Первая из них, расположенная непосредственно под мягкой мозговой оболочкой в самых верхних слоях коры, состоит из четырехугольных широких петель, лежащих в плоскости, параллельной наружной поверхности мозга. В слоях серого вещества соответственно размещению главным образом больших пирамидных клеток отмечается наличие капиллярной сети, сформированной из полигональных по форме петель, прилегающих плотно друг к другу.
При раздельной инъекции артерий мозга желатиной, окрашенной кармином, и вен желатиной, окрашенной синькой, Дюре получил заполнение обеих сетей серого вещества карминовой массой. Это обстоятельство дало ему повод высказать мнение о том, что обе капиллярные сети коры являются артериальными.
Часть венозной крови из артериальных капиллярных сетей коры оттекает в переходную или собственно капиллярную сеть, занимающую по своему положению последний миллиметр серого вещества. Расположенная на границе коры и белого вещества переходная сеть формируется в основном за счет деления коротких, или корковых, вен и при раздельной инъекции оказывается окрашенной в голубой цвет.
Артериальное снабжение белого вещества обеспечивается специальными «артериями белого вещества».
Последние представляют собой длинные артериальные стволы, которые при прохождении через кору отдают незначительное количество очень тонких нежных веточек и основную массу своих разветвлений сосредоточивают в белом веществе. Сформированная мельчайшими подразделениями ветвей длинных артерий артериальная капиллярная сеть белого вещества переходит в венозную сеть, в свою очередь вливающуюся в длинные вены. В соответствии с наличием артериальной и венозной сети в белом веществе в опытах Дюре здесь обнаруживались красная и голубая инъекционные массы.
Как можно видеть, Дюре создал довольно сложное и во многом неправильное представление о кровообращении в мозгу. Предложенное этим исследователем деление артерий мозга па короткие, ветвящиеся в коре или, самое большее, на границе ее с белым веществом, и длинные, доставляющие основную массу крови в белое вещество, предполагает не что иное, как раздельное артериальное снабжение серого и белого вещества мозга.
Более того, разграничение капиллярной сети коры на три слоя означает обособление артериальной части сосудистой сети от венозной. В самом деле переходная капиллярная сеть, располагающаяся в самом последнем слое серого вещества при переходе его в белое, по предположению Дюре, является местом сосредоточения продуктов обмена веществ всей массы клеточных элементов коры.
Другими словами, ток венозной крови из серого вещества направляется главным образом в переходную сеть. Из переходной сети основная масса крови идет по анастомозам, связующим ее с венозной сетью белого вещества, в крупные вены, которые в то же время представляют собой главные пути для оттока венозной крови из белого вещества в вену Галена.
Следовательно, согласно точке зрения Дюре, отток венозной крови из мозга в значительной мере происходит от его поверхности к желудочкам. Но при таких соотношениях клеточные элементы коры должны находиться в окружении одной артериальной крови. Этот вывод, вероятно, послужил для Дюре поводом к предположению о возможном возникновении вен в различных слоях коры, ломимо переходной сети, а также о частичном оттоке венозной крови из переходной сети через кору по направлению к венозной сети мягкой мозговой оболочки. Но и здесь Дюре не мог не оговориться, отметив, что ветвления коротких вен в се-
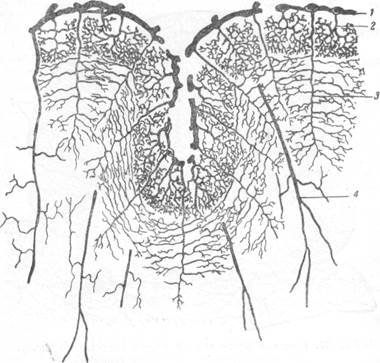
Рис. 72. Снабжение коры и белого вещества артериальной кровью
по Шарпи.
1 — артерии мягкой мозговой оболочки; 2 — артерии, снабжающие кору; 3 — средние артерии, снабжающие белое вещество; 4 — длинные артерии, снабжающие белое вещество.
ром веществе чрезвычайно скудны, и, следовательно, объем крови, оттекающий через толщу коры, очень невелик по сравнению с массой крови, устремляющейся в стволовые вены.
Несмотря на многие сомнительные данные, учение Дюре о коротких и длинных артериях мозговой субстанции было не только положено в основу анатомического распределения сосудов в мозгу его современниками, но признается и поддерживается рядом авторов и до настоящего времени. Как это иногда бывает в науке, некоторые положения учения Дюре в работах его последователей приобрели более крайнее выражение, чем это было высказано их основоположником. В качестве примера можно привести работу Шарпи (Charpy, 1899). Теоретические представления этого исследователя самым наглядным образом отражены на рис. 72, взятом из его работы. Из этого рисунка видно, что обособле-
ние зон артериального снабжения мозгового вещества выражено у Шар-пи значительно более резко, чем у Дюре.
Если Дюре считал, что короткие артерии коры дают боковые ветви не только в сером веществе, но и на его границе с белым, то Шарпи выделил корковые артерии в особую группу сосудов, принадлежащих со всеми их разветвлениями только коре. Вторая зона артериального снабжения, согласно точке зрения Шарпи, может быть отмечена в белом веществе, непосредственно подлежащем коре. Артерии, питающие этот отдел мозгового вещества, по его мнению, располагаются в коре между корковыми артериями и в сером веществе не дают боковых ветвей. Пер-
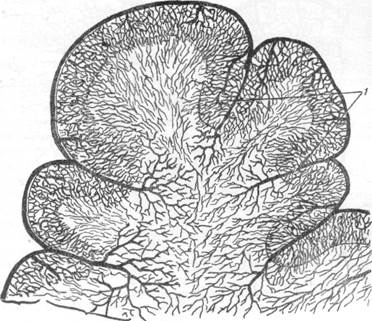
Рис. 73. Кровоснабжение коры и белого вещества по Якобу. 1 — артерии, снабжающие белое вещество.
вые ветви, отходящие под прямым углом к основному стволу, появляются только на границе серого и белого вещества. Глубокие слои белого вещества снабжаются третьей группой особенно длинных артериальных сосудов, разветвляющихся дихотомически.
Еще дальше пошел Якоб, утверждающий в своей монографии (1927), что длинные артерии, специально предназначенные для кровоснабжения белого вещества, погружаются в мозг только в глубине борозд и никогда не вступают в мозговое вещество с наружных поверхностей извилин (рис. 73).
Беркель (Berkel) и др. в своей работе, опубликованной в 1939 г., на основании изучения рентгенограмм сосудов мозга разделили артерии мозгового вещества на две группы.
Первая группа включает в свой состав корковые артерии (a. griseo-corticales) протяженностью в среднем до 2 мм. Эти короткие артерии идут параллельно друг другу, перпендикулярно к мозговой поверхности и имеют приблизительно одинаковую длину и объем. На срезе толщиной в 5 мм можно насчитать до 40 таких артерий на участке в 1 см длины.
Параллельно коротким артериям, и, следовательно, перпендикулярно к поверхности мозга по коре проходят длинные артерии, снабжающие белое вещество. Объединяемые в группу a. albo-corticales, длинные артерии меняют направление своего хода при достижении овального центра. Подойдя к овальному центру, такая артерия по прямой линии направляется от коры к соответствующему желудочку, где и оканчивается не доходя 1—1,5 мм до его боковой поверхности. Часто на некотором расстоянии от желудочка артерия распадается на две ветви, идущие параллельно друг другу по основному направлению ствола. Соответственно месту их отхождения от артерий мягкой мозговой оболочки a. albo-corticales имеют различную длину, достигая в лобной и затылочной доле 40 мм. На срезе толщиной в 5 мм располагается от 3 до 5 артерий на расстоянии 1 см.
Таковы данные ряда исследований различных авторов, придерживающихся точки зрения раздельного артериального снабжения серого и белого вещества головного мозга.
Почти одновременно с работой Дюре появилось иное представление о соотношении кровоснабжения в сером и белом веществе мозга.
Еще М. А. Тихомиров (1880) на основании изучения препаратов сосудистой сети мозга, налитой окрашенными массами, пришел к убеждению, что артерии, снабжающие белое вещество, отдают многочисленные ветви и в коре при своем прохождении через нее. Другими словами М. А. Тихомиров высказал впервые соображение о единстве кровоснабжения всей толщи стенки полушария.
Дальнейшее свое развитие эта мысль получила в работах Пфайфера.
В результате исследований большого материала Пфайфер пришел к отрицанию учения окоротких и длинных артериях и венах в том виде, как оно было предложено Дюре и некоторыми из его последователей. Это, однако, не означало, что артерии и вены мозга не могут различаться по длине их ствола, так как различные потребности клеточных скоплений в стенке полушария в кислороде и питательных веществах должны являться причиной существования в мозгу артерий и вен самой различной протяженности.
С точки зрения Пфайфера даже самые значительные по длине артерии или вены не предназначаются исключительно для кровоснабжения или оттока крови только белого вещества. Длинные артерии и вены при прохождении через кору своими ветвями участвуют в формировании капиллярной сети серого вещества, обеспечивая тем самым единство кровоснабжения белого вещества и коры.
Обзор литературы, опубликованной после появления работы М. А. Тихомирова и первой монографии Пфайфера, показывает, что последующее изучение вопроса о длинных и коротких артериях мозгового вещества не дало ничего нового по сравнению с тем, что было высказано уже этими исследователями.
В настоящее время нет никаких оснований возражать против предположения о том, что в мозговом веществе можно обнаружить артерии и вены самой разнообразной протяженности. Справедливость этого наглядно иллюстрируется прилагаемыми здесь микрофотографиями, взятыми из наших работ. Импрегнация сосудистой сети и в равной мере инъекция ее различными массами или прижизненная окраска сосудистой стенки убедительно свидетельствуют о том, что в мозгу имеются артерии, снабжающие только серое вещество, артерии, доставляющие кровь и в серое и белое вещество, и артерии, питающие преимущественно белое вещество.
Изучение многочисленных серий препаратов позволяет нам полностью присоединиться и несколько расширить разделение артерий стенки полушарий головного мозга, данное Кэмпбеллом в его работе, опубликованной в 1938 г. Рис. 61, а, б, в, г дает возможность убедиться в существовании в коре полушарий головного мозга артерий, основные ветви и ствол которых в своем распространении ограничиваются самыми верхними слоями коры. Средние слои серого вещества получают кровоснабжение по артериям со стволами большой длины по сравнению с предыдущими. Главная масса боковых ветвей этих артерий отходит от основного ствола в средних слоях, но ветвление той же артерии можно проследить и в более поверхностно расположенных слоях серого вещества.
Известная часть артерий доставляет кровь главным образом в нижние слои коры и в верхние слои подлежащего коре белого вещества. Соответственно основной области снабжения, основная масса наиболее мощных ветвей, как это видно из рис. 62, отходит от ствола артерии на уровне размещения нижних слоев клеток коры и верхних этажей белого вещества. Подобно артериям, основные области распределения ветвей которых ограничиваются верхними или средними слоями серого вещества, артерии последнего типа дают боковые ветви по всей длине ствола, начиная с верхних слоев коры.
Как можно видеть, выделение артерий в ту или иную группу в зависимости от места преимущественного отхождения боковых ветвей носит несколько искусственный характер, поскольку ветви от артерий самой различной длины отходят на всем протяжении хода сосуда.
Артерии, снабжающие белое вещество, должны быть отнесены к группе артериальных сосудов с наибольшей длиной основного ствола. Среди них в свою очередь можно различать артерии, погружающиеся в белое вещество на различную глубину. Общей характеристикой всей группы длинных артерий является более или менее слабое ветвление их в сером веществе, через которое они проходят по пути из мягкой мозговой оболочки. В том случае, когда артерия доставляет кровь в верхние или средние этажи белого вещества, боковые ветви различной мощности могут отходить от ее ствола на различных уровнях в сером веществе, а чаще всего в его нижних слоях. Артерия, подобно описываемой (рис. 62), уменьшается в объеме по мере своего хода и приходит в белое вещество с калибром значительно меньшим, чем тот, который она имела при от-хождении от артерии мягкой мозговой оболочки. Несколько иной вид имеют артерии, снабжающие самые глубокие слои белого вещества. Такая артерия обычно не дает боковых ветвей в сером веществе. Если же боковые ветви и отходят от основного ствола, то они имеют крайне незначительную длину и малый диаметр. Чаще всего первые ветви начинают отходить от ствола длинной артерии на том или ином уровне белого вещества, но так же, как и ветви артерий в коре, они незначительны по калибру. Благодаря тому, что длинная артерия на протяжении своего хода в коре и части белого вещества дает небольшое количество тонких ветвей, диаметр основного ствола ее меняется мало, даже тогда, когда артерия достигает самых глубоких слоев белого вещества. На уровне же этих слоев, т. е. в непосредственной близости от поверхности мозговых желудочков, от ствола длинной артерии отходят многочисленные ветви и образуют капиллярные сети.
Таким образом, длинные артерии, снабжающие самые глубокие слои белого вещества, занимают особое положение среди других артерий коры и белого вещества.
Все другие артерии посредством своих боковых ветвей и их дальнейших ветвлений вступают между собой в теснейшую связь по крайней мере посредством непрерывной капиллярной сети. Длинные же артерии, направляющиеся в глубокие околожелудочковые слои белого вещества, участвуют в формировании капиллярных сетей коры и верхних этажей белого вещества лишь в незначительной степени. Как уже говорилось, вся основная масса ветвей длинных артерий и возникающие от них капиллярные сети располагаются в глубине белого вещества в непосредственной близости от мозговых желудочков.
Дата добавления: 2015-05-16; просмотров: 1198;