Основные данные о строении артериальной сети мягкой мозговой оболочки у кролика 4 страница
Единство мозгового кровообращения, следовательно, осуществляется посредством капиллярного русла. Лишь очень редко в сосудисто-капиллярной сети мозгового вещества можно обнаружить анастомозы крупнее прекапиллярного размера, связывающие артерию с артерией и вену с веной вне капиллярного русла. При этом анастомозы чаще соединяют вену с веной, чем артерию с артерией (рис. 76, а). В полном соответствии с данными некоторых исследователей мы отмечаем, что анастомозы встречаются в венозной части сосудистой сети мозга чаще, чем в артериальной, и среди венозных сосудов белого вещества чаще, чем среди венозных сосудов коры.
На многих сериях препаратов, обработанных несколькими различными методами, выявляющими сосудисто-капиллярную сеть, мы никогда не наблюдали непосредственных связей между артерией и веной, т. е так называемых артерио-венозных анастомозов.
На основании результатов изучения большого материала мы пришли к выводу, что связи артерии с артерией и вены с веной крупнее прекапиллярного размера вне капиллярной сети должны быть отнесены к редким исключениям. Наличие их является результатом сохранения в том или ином участке сосудистой сети мозга эмбрионального строения, характерного для самого раннего этапа ее организации.
При разборе фактических данных, полученных при исследовании сосудистой сети мозга в процессе ее развития, мы уже видели, что определенные стадии развития дают нам возможность наблюдать совершенно особое расположение сосудов в стенке мозгового пузыря. Сосуды, прорастающие в матрикс, идут через поперечник мозгового пузыря параллельными стволами, соединяющимися между собой поперечными сосудами, почти такого же размера, как объединяемые ими стволы. Характерно, что преобразование сосудистого дерева и возникновение капиллярных сетей происходят именно там, где наличие нервных клеток вызывает наибольшую потребность в кислороде и питательных веществах. Так, например, на стадии восьмислойной стенки мозгового пузыря капиллярные сети располагаются там, где наблюдается наибольшее скопление клеточных элементов (в матриксе, коре, полосатых слоях). В других же слоях (в промежуточном и переходных), в которых сосудистая сеть в это время
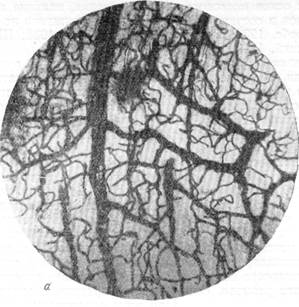
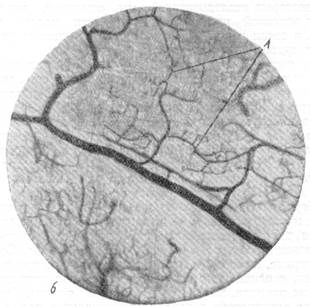
Рис. 76, а к б. Анастомозирование ветвей мозговых вен между собой в белом веществе и артерий в ретикулярной субстанции.
Импрегнация по методу В. И. Клосовского. Увеличение 100.
а — микрофотография с препарата мозга собаки, погибшей от острого отека через несколько часов после закрытия левой средней мозговой артерии. Представлено белое вещество области, расположенной между передней и средней мозговой артерией;
(анастомозирование вен);
б — анастомозирование ветвей артерий ретикулярной субстанции продолговатого мозга
между собой.
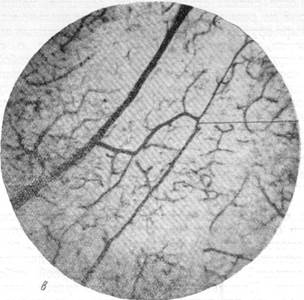
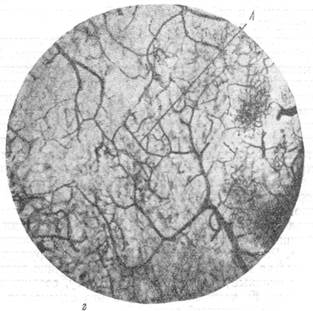
Рис. 76, в и г. Анастомозирование ветвей мозговых вен между собой в белом веществе и артерий ретикулярной субстанции.
в — анастомозирование ветвей артерий, вступающих в ретикулярную субстанцию продолговатого мозга с одной поверхности; г — анастомозирование ветвей артерий, вступающих в ретикулярную субстанцию продолговатого мозга с противоположных поверхностей. Микрофотографии б, в, г, сняты с препаратов продолговатого мозга, сосудистая система которого была инъицирована тушью. Увеличение 20. Анастомозы (А) указаны стрелкой.
находится на более ранних стадиях своего развития, капиллярные сети почти или совсем не развиты. В этих слоях параллельно следующие через стенку пузыря крупные сосуды соединены поперечными анастомозами.
Таким образом, непосредственная связь артерии с артерией или вены с веной сосудом крупного калибра представляет собой более примитивный тип соединения, чем капиллярная сеть. Если принять во внимание, что поздно оформляющие свою сосудисто-капиллярную сеть слои поперечника полушарий в окончательно сформированном мозгу соответствуют областям расположения белого вещества, то становится понятным, почему анастомозы в мозгу взрослого животного чаще встречаются в белом веществе, а не в коре.
Таким образом, изучение организации сосудисто-капиллярной сети мозга в онтогенезе определенно указывает, что анастомозы крупнее прекапиллярного размера представляют собой не что иное, как отражение эмбрионального характера строения сосудистой сети.
Интересно, что в составе сосудистой сети отделов головного мозга, филогенетически более старых, чем кора полушарий, можно отметить наличие анастомозов крупнее капиллярного размера. С подобного рода явлениями встретились Е. Н. Космарская и Е. Г. Балашева (1950) при изучении особенностей кровоснабжения ретикулярной субстанции продолговатого мозга. Просматривая серии препаратов мозга кошек и собак, сосудистая сеть которых была инъицирована тушью, трипановой синью или импрегнирована серебром по методу Б. Н. Клосовского, эти авторы установили, что артерии ретикулярной субстанции широко анасто-мозируют между собой. Анастомозы различного калибра, по своей величине в несколько раз превосходящие размер прекапилляра, соединяют ветви как одной и той же (рис. 76, б), так и различных артерий между собой (рис. 76, в, г).
В тех случаях, когда подлинные анастомозы соединяют ветви различных артерий между собой, можно различать два типа. В первом из них, как это можно видеть из рис. 76, в, анастомозом связаны ветви двух артерий, вступающих в вещество продолговатого мозга с какой-либо одной поверхности и идущих поэтому друг возле друга в одном направлении. Во втором типе соединенными анастомозами оказываются ветви артерий, вступающие в продолговатый мозг с разных поверхностей его и направляющихся, следовательно, навстречу друг другу. Так, на рис. 76, г, приведенном для иллюстрации вышесказанного, можно видеть большое количество анастомозов между ветвями двух крупных артерий. Одна из них вступила в вещество продолговатого мозга с вентральной, другая — с дорзальной поверхности его.
Характерно, что соединение сосудов крупными анастомозами встречается и в патологических случаях. С подобными явлениями мы столкнулись при изучении сосудистой сети мозга людей, при жизни страдавших болезнью Дуана. Рис. 77 показывает своеобразную структуру сосудистой сети серого вещества в случаях болезни Дуана. Можно видеть, что сосуды коры, расположенные параллельно друг другу, соединяются поперечными анастомозами по тому же типу, как это отмечается на ранних стадиях организации сосудистой сети.
Мы пока не имеем возможности различать артерии и вены среди сосудов, врастающих в стенку мозгового пузыря на самых ранних стадиях развития. Предпринятые в этом направлении работы, безусловно, прольют свет на многие неясные сейчас стороны мозгового кровообращения и, в частности, выясняет характер взаимодействия артериальной и венозной сосудистой сети на самых первых этапах ее организации.
Вполне возможно, что на этих этапах развития может иметь место непосредственное соединение артерий с венами, обеспечивающее быстрый отток крови, содержащей продукты обмена веществ клеток матрикса, но изучение этих самых ранних стадий развития и является наиболее технически трудным. К сожалению, в настоящее время мы не могли приготовить препараты, рассмотрение которых позволило бы проследить эти ранние стадии организации сосудисто-капиллярной сети в мозгу, так как на имеющихся в нашем распоряжении препаратах представлены не толь-
|
Рис. 77. Структура сосудисто-капиллярной сети в
коре при болезни, Дауна.
Микрофотография с препарата, окрашенного по методу Эроса. Увеличение 100.
ко артерии и вены матрикса, но и объединяющая их капиллярная сеть. Поэтому следует признать, что имеющиеся данные о развитии сосудисто-капиллярной сети в онтогенезе пока недостаточны для того, чтобы представить себе возможность существования артерио-венозных анастомозов в мозгу взрослого животного. Непосредственные связи артерий и вен между собой, если они встречаются, должны рассматриваться как чрезвычайно редкая аномалия строения сосудистой сети мозга.
Отсутствие в мозгу взрослого животного артерио-венозных анастомозов подтверждают также опыты с раздельной инъекцией артериальной и венозной части сосудистого русла мозговой ткани. Раздельная инъекция только потому и возможна, что в нормальных условиях не существует непосредственного тока крови из артерий в вены, помимо капиллярной
сети.
Трудно также представить себе наличие артерио-венозных анастомозов в мозгу взрослого животного и с физиологической точки зрения. Действительно, в других органах с постоянно меняющейся циркуляцией крови, в зависимости от потребностей ткани или всего органа в целом в каждый данный момент, артерио-венозные анастомозы могут играть опре-
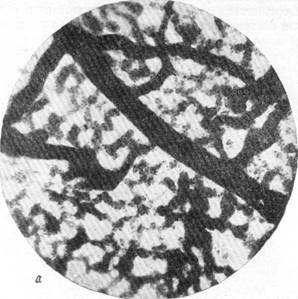
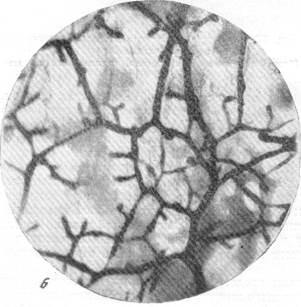
Рис. 78. Сравнительная диференцировка артериальной и венозной сети мягкой мозговой оболочки на различных стадиях онтогенетического развития человека. а — венозная сеть плода длиной 30 см; б — артериальная сеть плода длиной 30 см

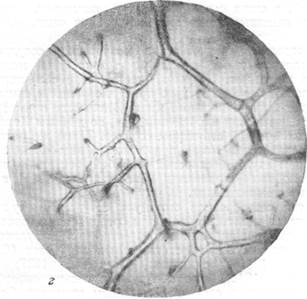
Рис. 78. Сравнительная диференцировка артериальной и венозной сети мягкой мозговой оболочки на различных стадиях онтогенетического развития человека.
в — венозная сеть плода длиной 36 см; г — артерильная сеть плода длиной 36 см, Микрофотографии с гиперемированной мягкой мозговой оболочкой плодов, окрашенной
по методу Эроса. Увеличение 50,
деленную роль в кровообращении [Е. Кларк (E.Clark), 1938]. В мозговой же ткани процессы жизнедеятельности требуют постоянного тока крови, при котором существование артерио-венозных анастомозов утрачивает всякий смысл. При наличии их в сосудистой сети мозга они являлись бы обходным путем для крови, благодаря чему кровь не заполняла бы всего капиллярного русла. Между тем нервные клетки получают кислород и питательные вещества из крови, циркулирующей именно по капиллярам, и таким образом нормальная жизнедеятельность нервных клеток зависит от постоянства тока крови в капиллярном русле. Следовательно, наличие артерио-венозных анастомозов в сосудистой сети мозга должно было бы нарушить постоянство тока крови в сосудистой сети и тем самым служить причиной нарушений обмена в нервных клетках.
Выше мы уже отмечали, что артерии мозга по сравнению с венами являются сосудами более диференцированными. Артерии меньше ветвятся, чем вены. Основная масса крупных боковых ветвей артерий обычно сосредоточивается на определенном уровне, тогда как крупные ветви могут вливаться в вену по всему ее ходу. Благодаря этому обстоятельству область мозгового вещества охватывается веной в большей степени, чем снабжающая эту область артерия. Большая область охвата и большая густота венозной капиллярной сети создают большие возможности перемещения крови в ней по сравнению с перемещением крови по артериальной капиллярной сети.
Более близкое к эмбриональному строению обнаруживает и венозная сеть мягкой мозговой оболочки. На рис. 78 для сравнения приведены микрофотографии венозной и артериальной сети мягкой мозговой оболочки у плодов человека 30 и 36 см длины. Эти микрофотографии подтверждают предположение о сохранении венозной сетью мозга на каждой стадии развития более примитивного строения по сравнению с артериальной, в связи с чем становится понятным, почему анастомозы между венозными стволами встречаются чаще, чем между артериями.
Тип распределения сосудов в виде непрерывной сосудисто-капиллярной сети является характерным не только для мозга человека и лабораторных животных. Подобное же строение сосудистой системы в мозгу наблюдается у животных, принадлежащих к самым различным классам и даже типам животного мира. В качестве примера можно указать на мозговые ганглии каракатицы, где, по данным Кахаля, отмечается наличие сетеобразной структуры сосудов. Такое же расположение сосудов обнаруживается в мозгу примитивных позвоночных миксин, рыб (за исключением акул), бесхвостых амфибий, рептилий (исключая ящериц), однопроходных [Сандерланд (Sunderland), 1941], а также всех плацентарных млекопитающих.
Вместе с тем сетеобразная структура сосудов в мозгу животных и человека не является единственным типом организации кровоснабжения его.
При исследовании анатомического строения сосудистой системы в мозгу lamprey, petromyzon, хвостатых амфибий [Крэги (Cralgie), 1940], ящерц (Крэги, 1941), сумчатых (Вислоки, 1937, 1939; Крэги, 1938; Сандерланд, 1941) было обнаружено весьма своеобразное и характерное расположение сосудов в мозговом веществе. Особенно отчетливо выраженным и типичным оно оказалось в мозгу кенгуру и опоссума, которые и подверглись наиболее тщательному изучению.
Характерной особенностью кровоснабжения опоссума (так же как и всех других животных, относящихся к этому типу) является расположение сосудов во всех органах тела и в том числе в мягкой мозговой обо-
лочке в виде непрерывной сети. Однако сосуды, проникающие из мягкой мозговой оболочки в мозговое вещество, имеют уже совершенно своеобразный характер.
Мы уже указывали, что у человека и животных с типичной для них сетеобразной структурой сосудов в мозгу нельзя отметить какой-либо закономерности в порядке вхождения артерий в мозговое вещество. Артерии проникают в мозг то совсем рядом друг с другом, то отделены одна от другой большим расстоянием. Никогда не удается проследить, чтобы артерия и вена имели вход и выход из мозговой субстанции в одном участке. В мозговом веществе артерии также, как правило, никогда не сопровождаются венами, а капиллярные сети той и другой обнаруживают сложное взаиморасположение.
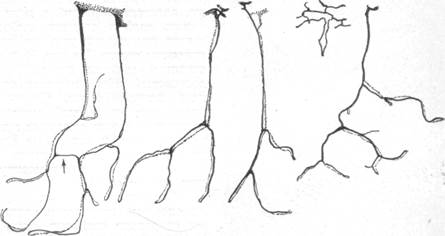
Рис. 79. Различные по форме конечные артерия в мозгу опоссума (по Вислоки).
Иные соотношения наблюдаются в мозгу опоссума. Участок поверхности мозга, в котором артерия проникает в мозговое вещество, располагается в непосредственной близости с участком, на котором из мозга выходит соответствующая данной артерии вена. В мозговом веществе артерия и вена идут друг возле друга, причем каждой ветви артерии вплоть до капилляров точно соответствует такая же по форме ветвь вены. На рис. 79, взятом нами из работы Вислоки (1939), показаны различные по сложности парные конечные сосуды, располагающиеся в мозгу опоссума.
Отсутствие связей между каждой такой парой сосудов превращает каждую из них в анатомическую и функциональную единицу. Таким образом, здесь имеется настоящая конечная артерия, закупорка которой будет иметь следствием выпадение мозгового вещества во всей области распределения ее разветвлений. Для характеристики расположения в мозгу опоссума конечных артерий и характера выпадения ткани при закрытии просвета какой-либо артерии интересны эксперименты Шаррера (1939).
Через 48 часов после инъекции спор ликоподия в сонную артерию опоссума Шаррер наблюдал в зернистом слое мозжечка распад и исчезновение нервных клеток, располагавшихся в радиусе 25 м от закрытой артерии.
На рис. 80 видно, что каждая артерия мозгового вещества опоссума обеспечивает кислородом и питательными веществами вполне определенную область. В соответствии с этим одна пара сосудов в мозгу опоссума располагается от другой на равном расстоянии, не превышающем 50 м. в совершенно правильном порядке.
|
Таким образом, в мозгу опоссума и других животных, мозг которых
снабжается кровью также по сосудам конечного характера, суще
ствует ограниченное количество
капилляров, необходимое для
жизнедеятельности мозговой
гкани.
Характерно, что, несмотря на сетеобразное распределение артерий на поверхности мозга опоссума, врастание артерий в мозг на самых ранних стадиях эмбриогенеза происходит уже по типу конечных петель. На рис. 81 представлено врастание артерий в продолговатый мозг эмбриона опоссума длиной 14 мм. Отчетливо видно, как из мягкой мозговой оболочки в стенке мозговых пузырей прорастают отдельные сосудистые единицы, совершенно обособленные, не анастомозирую-щие между собой.
Снабжение мозгового вещества опоссума конечными артериями сохраняет свой тип даже в тех случаях, когда в мозг названного животного имплантируется участок мозга какого-либо другого животного, для которого характерно сетеобразное распределение сосудов в мозгу.
| Рис. 80. Характер снабжения артериальной кровыо мозгового (вещества опоссума (по Шарреру). |
Такого рода эксперимент был проделан в свое время Шаррером (1939). При имплантации ib мозг опоссума участка мозговой ткани крысы или морской свивки Шар-рер (1939) отметил, что регенерирующие сосуды мозга опоссума врастали в пересаженный участок по типу конечных. В тех же случаях, когда участок мозга опоссума пересаживался в мозг морской свинки, сосудистая сеть имплантата приобретала сетеобразное строение, характерное для мозга морской свинки.
Наблюдения Шаррера пока не нашли еще дальнейшего развития в работах других исследователей. Решение вопроса о факторах, определяющих характер строения сосудистой сети мозгового вещества, имеет большое значение. Данные, полученные при решении этого вопроса, мог-ли бы объяснить, почему сетеобразная структура сосудов встречается не только у позвоночных, но и у ряда беспозвоночных, и, наоборот, почему конечные артерии могут быть найдены не только у беспозвоночных, но в различных группах позвоночных животных.
Было бы неправильно думать, что в высокодиференцированноммозгу человека существует только один тип распределения сосудов в виде сети.
Так, ворсинки сосудистых сплетений получают кровь по сосудам, несколько напоминающим петли. Совершенно особое строение сосудистой сети имеет также area postrema.
При обработке всего мозга животного или человека предложенным нами методом импрегнации сосуды area postrema не выявляются подобно сосудам plexus chorioideus. Кроме того, эндотелий капилляров как того, так и другого образования одинаковым образом относится к введению в сосудистое русло витальных красок.
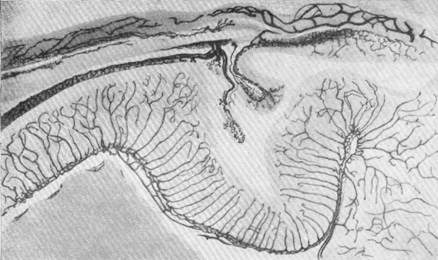
Рис. 81. Врастание сосудов в стенку мозговых пузырей у эмбриона опоссума длиной 14 мм (по Вислоки).
Известно, что в мозгу взрослых особей эндотелий капилляров в силу особого химического состава представляет собой барьер для витальных красок. Благодаря этому краска не выходит за пределы капиллярного русла и окрашивание мозговой ткани не происходит. Эндотелий же капилляров сосудистых сплетений и area postrema пропускает витальные краски.
При изучении гистологических препаратов можно также отметить своеобразное, отличающееся от других участков мозга, расположение сосудов в описываемом поле. Артерии и вены входят в area postrema друг возле друга и в дальнейшем своем ходе обнаруживают сложные взаимоотношения. Создается впечатление, что один сосуд как бы обвивается вокруг другого.
Подобные же взаимоотношения между артерией и веной при конечном характере их наблюдаются в массивных хориопапилломах (рис. 82, а), а также были найдены нами в случае мультиформной спон-гиобластомы, локализовавшейся в полушариях головного мозга (Б. Н. Клосовский, Е. Н. Космарская) (рис. 82, б). То же удается отметить и на приведенных выше рисунках из работ Вислоки и Шаррера (рис. 79, 81), хотя этот факт указанными исследователями отмечен не
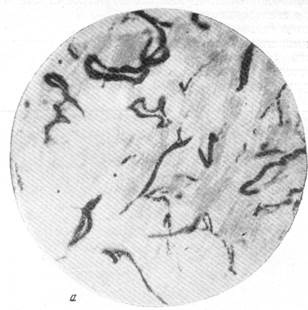
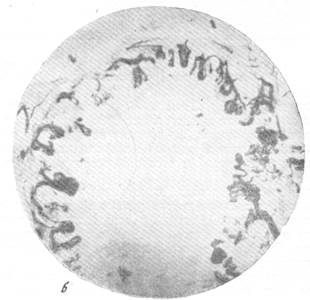
Рис. 82.
а — характер сосудов в хориопапилломе. Обращает на себя внимание организация капилляров в виде конечных петель. Окраска по методу Эроса. Увеличение 100. — характер сосудов в мультиформной спонгиобластоме. а—расположение сосудов в виде конечных петель в стенке кисты. Увеличение 100.
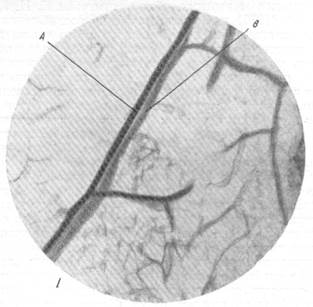
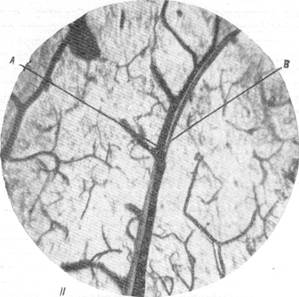
Рис. 83.
I, II — взаимное расположение артерий и вен в продолговатом мозгу млекопитающих. А — артерия; В — вена. Инъекция сосудистой сети тушью. Увеличение 20.
был. Как будет видно в дальнейшем, соотношение артерии и вены в случаях снабжения мозга конечными сосудами (у опоссума) играет существенную роль в понимании процессов питания и отдачи продуктов обмена веществ нервными клетками, располагающимися вокруг них.
Соотношения артерий и вен, несколько напоминающее соотношения их в мозгу сумчатых животных, были отмечены Е. Н. Космарской и Е. Г. Балашевой (1950) при исследовании кровоснабжения продолговатого мозга млекопитающих. Изучение срезов мозга кошки и собаки, сосудистая сеть которых была инъицирована тушью, трипановой синью или импрегнирована серебром, дало возможность констатировать, что для артерий и вен продолговатого мозга является типичным вхождение первой в мозг и выхождение второй из него в непосредственной близости в одном и том же участке мягкой мозговой оболочки. Это в равной мере относится как к срединным, так и к боковым артериям и венам. В дальнейшем ходе артерий и вен в веществе продолговатого мозга могут обнаруживаться два типа взаимоотношений. В большинстве случаев основной ствол артерии не только сопровождается основным стволом вены, но артерия и вена закручиваются друг возле друга так, как это можно видеть на рис. 83, /, //. Иначе говоря, мы наблюдаем здесь то же взаимное расположение артерий и вен, как в мозгу сумчатых, имеющих конечные артерии. Однако сходство ограничивается только соотношением основных стволов, тогда как разветвления артерий сопровождаются соответствующими разветвлениями вен далеко не всегда.
В продолговатом мозгу млекопитающих животных лишь в редких случаях бассейн снабжения боковой или срединной артерии совпадает с бассейном, с которого собирает кровь вена, идущая рядом с артерией. Обычно основной ствол артерии имеет большую протяженность, чем сопровождающий его ствол вены. Иными словами, область снабжения данной артерии не совпадает с областью, с которой собирает кровь соответствующая ей вена.
В некоторых случаях удается отметить обратное явление, когда ствол вены имеет большую протяженность по сравнению со стволом ар-терии. Соответственно область, с которой собирает кровь вена, здесь располагается дальше области снабжения рядом идущей артерии.
Довольно часто можно наблюдать, что артерия и вена на значительном расстоянии идут друг возле друга или закручиваются одна вокруг другой. Однако на некотором уровне стволы артерии и вены расходятся, причем артерия направляется в одну сторону, а вена в другую. Следовательно, и в этих случаях область мозгового вещества, получающая кровь по данной артерии, территориально не совпадает с областью, с которой собирает кровь соответствующая вена.
Таким образом, в картине хода артерий и вен в веществе продолговатого мозга млекопитающих мы отмечаем лишь только некоторое сходство с ходом артерий и вен в мозгу сумчатых животных.
Импрегнация всей сосудисто-капиллярной сети мозга по методу Б. Н. Клосовского позволяет отмстить, что в сетеобразной структуре сосудов, характерной для полушарий головного мозга человека и большинства позвоночных животных, существует все же ясно выраженная обособленность ядерных образований. Мы уже указывали на снабжение подкорковых ядер особыми артериями, распадающимися в них на капиллярную сеть, анастомозирующую с капиллярной сетью окружающего белого вещества лишь в капиллярном русле. Подобные же соотношения имеют место при снабжении кровью отдельных ядерных образований в различных отделах головного мозга человека и млекопитающих.
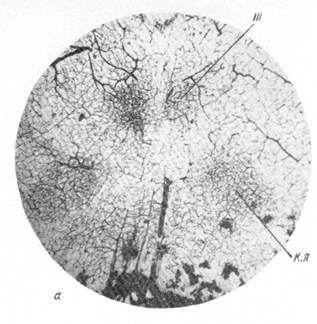
Рис. 84. Сосудисто-капиллярная сеть различных ядер и взаимодействие ее с капиллярной сетью окружающего мозгового вещества.
а — срез через средний мозг; III —ядро глазодвигательного нерва; к. я. — красное ядро (увеличение 10); б— ядро блоковидного нерва (увеличение 64).

Рис. 84. Сосудисто-капиллярная сеть различных ядер и взаимодействие ее с капиллярной сетью окружающего мозгового вещества. в — верхняя олива (увеличенние 64); г — срез через продолговатый мозг; х — ядро.
блуждающего нерва (увеличение 10). Импрегнация серебром по методу Б. Н. Клосовского.
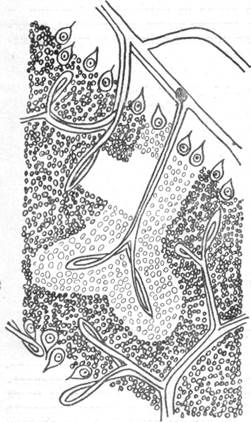
На рис. 84, а, взятом из нашей работы, на срезе, сделанном через средний мозг, представлена сосудисто-капиллярная сеть. Видно, что как ядро глазодвигательного нерва, так и красное ядро обладают капиллярной сетью, настолько резко обособленной от капиллярной сети окружающего мозгового вещества, что вполне возможно выделить местоположение того и другого образования. Те же соотношения можно видеть и на рис. 84, б, где отчетливо выделяется капиллярная сеть в области ядра блокового нерва, или на рис. 84, в, представляющем верхнюю оливу, или на рис. 84, г, на котором не менее отчетливо выделяется среди окружающей капиллярной сети капиллярная сеть ядра блуждающего нерва (образующаяся в результате ветвления специально идущей к данному ядру артерии).
Повсюду, как правило, можно отметить очень слабое взаимодействие капиллярных сетей различных ядер с капиллярной сетью остального мозгового вещества, обособление их и отсутствие подлинных анастомозов.
Выше мы подробно разобрали, что непрерывность сосудистой сети серого и белого вещества каждого в отдельности и вместе взятых осуществляется только в капиллярном русле. Капилляры, общая протяженность которых по некоторым данным [Линдгрин (Lindgreen), 1940, и др.] достигает 110 км, создают непрерывную структуру сосудистой сети в мозгу. Именно благодаря отсутствию в коре и белом веществе подлинных анастомозов артерии мозга оказываются конечными в функциональном отношении не только в патологических, но и в нормальных условиях. Иными словами, кровь, поступающая в серое вещество коры по радиальным артериям, выводится на периферию мозга, преимущественно по соответствующим коротким венам серого вещества. Кровь, направляюща-ся в белое вещество по длинным артериям, выводится на поверхность, преимущественно по соответствующим венам белого вещества. Таким образом, несмотря на существование в мозгу густой непрерывной капиллярной сети, кровь циркулирует в мозгу как бы по артерио-венозным единицам, представляющим собой наиболее короткие пути для тока крови в мозгу.
Конечно, эти артерио-венозные единицы не являются анатомически фиксированными частями сосудистой сети, как это наблюдается в мозгу сумчатых животных (кенгуру), а создаются в зависимости от функциональной деятельности мозговой субстанции.
В связи с изложенным становятся понятными данные о скорости тока крови в мозгу человека в нормальных условиях, полученные при введении контрастных веществ в ток крови общей сонной (внутренней сонной) артерии. Рентгенологические исследования показывают, что контрастные вещества протекают с током крови через систему внутренней сонной артерии в течение 2 секунд, в то время как по системе наружной сонной артерии контрастное вещество проходит от 6 секунд до 5 минут (Мониц, 1940, М. Д. Гальперин, 1950, М. Б. Копылов, 1950). Быстрота тока крови по системе внутренней сонной артерии может быть объяснена только наличием в мозгу коротких путей — арте-рио-венозных единиц.
Глава IV О КОЛЛАТЕРАЛЬНОМ КРОВООБРАЩЕНИИ В МОЗГУ
Как отмечалось выше, анатомо-гистологические данные указывают на распределение сосудов в мягкой мозговой оболочке и внутри мозгового вещества в виде более или менее сложно построенных сетей. При этом непрерывность сосудистой сети в мягкой мозговой оболочке обеспечивается наличием сосудов-анастомозов разнообразного размера. В соответствующем разделе подробно описывались сосуды-анастомозы, соединяющие в смежных зонах кровоснабжения крупные ветви основных мозговых артерий между собой. Существование такого рода анастомозов может обеспечить широкое перемещение крови из области распределения ветвей одной основной мозговой артерии в область другой при закупорке какой-либо из них. Вместе с тем анастомозы соединяют между собой также ветви одной и той же артерии в области распределения ветвей любой основной мозговой артерии. Наличие этих анастомозов при нарушении нормальной циркуляции крови на территории распределения ветвей одной из основных мозговых артерий создает возможность передвижения крови из одного участка мозговой поверхности в другой. Основой же непрерывности сосудов в мозговом веществе является капиллярная сеть. Таким образом, возможно перемещение крови из одного отдела мозга в другой при выключении артерии, питающей ту или иную область в нормальных условиях. Иначе говоря, анатомическое строение сосудистой сети в мягкой мозговой оболочке и внутри мозгового вещества допускает переключение питания мозгового вещества в бассейне закрытой артерии на кровоснабжение по коллатералям.
Дата добавления: 2015-05-16; просмотров: 1089;