Животные со слабо развитыми или редуцированными внутренними сонными артериями и хорошо развитыми позвоночными артериями
Грызуны (зайцы, белки, морские свинки)
Непарнокопытные (лошадь)
| Слабо развита, редуцирована | Развита |
| Слабо | Развита хо- |
| развита | рошо |
Нет
То же
Нет
То жег
Животные со слабо развитыми внутренними сонными и позвоночными
артериями
Хищные (собака)
Развита несколько слабее, чем у медведя
Развита слабо
Есть
Животные с редуцированными внутренними сонными и редуцированными или слабо» развитыми позвоночными артериями
Хищные (все представители семейства кошек, гиена полосатая)
Парнокопытные (крупный рогатый скот: овцы, олени, лани)
Китообразные (дельфины)
| Редуцирована То же | Развита слабо То же |
| » » | Редуцирована |
Есть
Есть
щимися в мозговое вещество, образуют артериальную часть круга кровообращения полушарий головного мозга.
Второй круг кровообращения формируется артериями, питающими, подкорковые образования. Эти артерии, которые по старой номенклатуре-называются сосудами центрального круга, обладают рядом особенностей. Одна из этих особенностей заключается в том, что большинство сосудов. отходит от главных стволов на основании мозга почти под прямым углом (Гюбнер, М. A. Тихомиров, Г. Д. Аронович и др.).
После отхождения от передней, средней или каких-либо других: артерий эти сосуды поступают в вещество мозга и прямо направляются к снабжаемой ими области. На всем пути от места отхождепия до снабжаемого ими района артерии имеют одинаковый продает и почти-не ветвятся. Лишь вступив в область васкуляризацпи, они отдают большее количество мелких ветвей.
Следует указать также, что подавляющее большинство артерий,. снабжающих подкорковые образования, не анастомозирует между собой и связано друг c другом только капиллярной сетью. Это обстоятельство отмечено многими исследователями не только 'в отношении мозга человека, но и мозга животных (Гюбнер, М. А. Тихомиров, Эбби, 1934; Кэмпбелл, 1938, и др.).
Ряд авторов делит сосуды для подкорковых образований, отходящие от средней мозговой артерии в количестве 5—8, на две группы: внутренние и наружные (M. А. Тихомиров, Б. К. Гиндце я др.).
Внутренние ветви обеспечивают кровью переднюю часть бледного» шара (globus pallidus) и внутреннюю капсулу (capsula interna). Наружные ветви, более длинные и (большего (калибра, распределяются в шорлу-пе (putamen) и в средней части хвостатого ядра (n. caudatus).
Среди ветвей передней мозговой артерии, снабжающих кровью подкормовые образования, особого вниманий заслуживает артерия, получившая название гюбнеровской, по имени впервые описавшего ее исследователя (рис. 1,3).
Согласно результатам наблюдений Айткена (Aitken, 1909), в 80%; случаев эта артерия берет свое начало от базального участка передней-мозговой артерии, но иногда может возникать я от внутренней сонной, артерии в месте деления ее на основные ветви и даже от средней мозговой артерии. В одном из своих случаев Критчли наблюдал отхождение гюбнеровекой артерии от передней мозговой уже после возникновения передней соединительной артерии.
Мы в свою очередь неоднократно отмечали подобный же способ отхождения тюбнеровской артерии на мозгу людей, погибших от различных опухолей мозга.
Этот сосуд, являющийся, по данным Эбби (1934), остатком анастомозов, (проходивших у низших животных через древний обонятельный мозг и вокруг него, не упоминается даже в работах, посвященных вопросам васкуляризации мозга. Примером в этом отношении может служить последняя, уже. неоднократно упоминавшаяся, работа, Б. К. Гиндце (1947). Подобно ряду исследователей, Б. К. Гиндце не выделяет гюбне-ровскую артерию среди мелких, как он называет, «артериол» передней мозговой артерии.
1 Существуют некоторые отличия в сообщениях различных исследователей отно-сительно количества, способа отхождения и областей васкуляризации отдельных ветвей, Интересующиеся подробностями найдут их в работах вышеупомянутых авторов, а также у Фуа, Бонне (1926), Эбби (1934) и др.
Между тем эта весьма важная в физиологическом отношении артерия представляет собой довольно длинный ствол с диаметром до 1,5 мм. От места своего отхождения (гюбнеровская артерия направляется назад к переднему продырявленному веществу (проходя, таким образам, расстояние около 2,5 см) и погружается в мозговое вещество в области передней продырявленной пластинки.
Внутри мовга гюбнеровская артерия ветвится на три ствола и распределяется ib голоске хвостатого ядра, в передней трети скорлупы, в наружном сегменте бледного шара и в переднем колене внутренней капсулы. Оказания о вариациях в размере, месте отхождения и способе ветвления этой артерии можно найти в работе А. Я- Подгорной (1936), выполненной в нашей лаборатории.
От задней соединительной артерии отходит несколько мелких сосу-дов (6—8 по М. А. Тихомирову и Б. К. Гиндце, б по Дюре), веточки которых распределяются -в зрительном бугре, питают зрительный перекрест, зрительный тракт, титечные тела, серый бугор, воронку и наружную часть ножек мозга.
Эти образования, за исключением зрительного тракта и перекреста, получают кровь таижа от базальныж ветвей задней мозговой артерии, снабжающих, «роме того, большую часть гипоталамической области, шишковидную железу, сосудистые сплетения третьего и боковых желудочков. BieTBin от ствола задави мозговой артерии или от одной из ее периферических ветвей доставляют кровь к передним буграм четверохолмия (corpus quadrigeminus), в то время как задние бугры снабжаются кровью по верхней мозжечковой артерии. От имаста возникновения задней мювгеной артерии, а та[КЖ'в и от1 самой этой артерии отходят сосуды, питающие (красное' ядро.
В настоящее время исследователи насчитывают при артерии, снабжающие это образование.
Две из них возникают у места отхожденяя задней Мозговой артерии от основной и погружаются в заднее продырявленное вещество (soibstantia peirforata posterior).
Одна из этих артерий идет в восходящем направлении вблизи от средней линии и возле ядра глазодвигательного нерва делится на две ветви. Первая веточка вступает в красное ядро, вторая снабжает ядро III пары чфепномозрфых нервов.
Вторая артерия в .свою очередь делится на две ветви: одну, для красного ядра, и другую, распределяющуюся в околожелудочковых отделах зрительного бугра.
Третья артерия красного ядра берет начало обычно от задней мозговой артерии, а в некоторых случаях и от верхней мозжечковой артерии, в том участке, где эта сосуды огибают ножку мозга.
Соответственно трем описанным артериям, участвующим в кровоснабжении красного ядра, различают три типа патологических синдромов его при закупорках питающих сосудов,
Сосудистые сплетения боковых жачудочков получают кровь, с одной стороны, от передней артерии сосудистого сплетения, являющейся ветвью внутренней сонной артерии, а с Другой—по одной из ветвей задней артерии сосудистого сплетения, отходящей от ствола задней мозговой артерии. Сосудистое сплетение третьего- желудочка снабжается также из двух источников. Основная масса крови в сосудистое сплетение третьего желудочка поступает по .ветви задней артерии сосудистого сплетения, но часть артериальнойкрови доставляется ветвью передней артерии сосудистого сплетения.
Передняя артерия сосудистого сплетения (a. chorioidea anterior) представляет собой сосуд незначительного калибра, отходящий обычно от внутренней сонной артерии одним или двумя стволами. Нередки случаи, когда эта артерия отходит от средней мозговой артерии, а иногда даже и от задней соединительной артерии. Помимо сосудистых сплетений
|
бокового итретьего желудочков, ветви описываемой артерии снабжают заднюю часть бледного шара, передние и средние отделы хвостатого ядра и скорлупы, ножки мозга, зрительный тракт, наружные передние части зрительного бугра, аммонов рог. Каждая из артерий сосудистого сплетения делится на две группы ветвей. Одна группа питает стенки желудочков, другая — вновь распадается на еще более мелкие ветви, образующие артерии в сосочках сплетения.
| Рис. 8. Аномальные формы основной и позвоночной артерии (по Стопфорду). |
Основным источником кровоснабжения зрительного бугра является задняя мозговая артерия. Но, помимо ветвей этой артерии, в зрительном бугре распределяются также ветви средней мозговой и задней соединительной артерий.
Кроме тола, наружная передняя часть зрительного бугра получает кровь по передней артерии сосудистого сплетения.
Гипоталамическая область получает кровоснабжение по нескольким мозговым артериям. Передняя часть гипоталамуса снабжается ветвями передней мозговой артерии, в боковых его отделах распределяются ветви
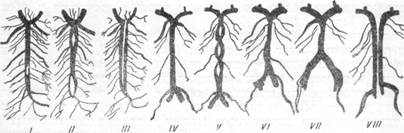
Рис. 9. Различные виды основной артерии у человека и животных
(пo Гиндце).
внутренней сонной и задней соединительной артерий, задняя часть питается от задней мозговой и задней соединительной артерии. В снабжении артериальной кровью аммонова рота принимают участие в основном тве мозговые артерии: передняя артерия сосудистого сплетения, дающая ветви к задней части рога, и задняя мозговая артерия.
Кровобращение моста, мозжечка и продолговатого мозга в свою очередь может быть выделено в самостоятельный круг. Основным источником, обеспечивающим кровоснабжение задней части мозга (мозжечка, варолиева моста к продолговатого мозга), являются парные позвоночные артерии.
На основании своего материала Стопфорд сообщает, что только в 8% обследованных ям случаев диаметр правой позвоночной артерии был равен диаметру соответствующей артерии слева. Таким образом, приведенные данные свидетельствуют, что в подавляющем большинстве случаев отмечается неравномерность диаметров позвоночных артерий. По наблюдениям того же автора, в большом проценте случаев неравномерность эта выражена крайне резко: так, в 22% изученных им мозгов просвет позвоночной артерии одной стороны превышал просвет соответствующей артерии другой стороны в два раза (рис. 8). На значительную частоту случаев неравномерности калибра позвоночных артерий
указывает также С. М. Огнева (1944) в своей работе, посвященной исследованию ваакуляризации варолиева моста, и продолговатого моага человека. Отмечено это и нами при изучении опухолевого материала
При входе в полость черепа позвоночные артерии, подобно внутренним сонным артериям, образуют изгибы, отчетливо видимые на рис. 2.
В полости черта позвоночные артерии сливаются в непарную основную артерию, располагающуюся в основной борозде на основании мовга. Исследуя развитие описываемых артерий у человека, Шмайдель (1937) пришел к выводу, что образование основной артерии происходит благодаря слиянию двух параллельно идущих позвоночных артерий. В отношении времени, к которому можно было бы отнести начало преобразования передних отделов позвоночных артерий в непарную основную, данные различных исследователей несиолько расходятся.
Так, Конгдон (Oongdon, 1922) считает, что слияние обеих артерий начинается еще тогда, когда артерии неполностью сформированы по Всей своей длине и расположены не вполне симметрично. Шмайдель и Гохштеттер относят этот процесс ik тому этапу развития, на котором позвоночные артерии вполне оформились как сосуды.
Отсутствие слияния позвоночных артерий, располагающихся в полости черепа в виде двух параллельных стволов (рис. 9, VIII), у человека наблюдается крайне редко, ihoi довольно часто (встречается у обезьяны ателес [Б. К. Гиндце, 1947].
В большинстве случаев слияние позвоночных артерий имеет место не на всем протяжении. Этот вид слияния (рис. 9, /, //, ///, V) характеризуется наличием «островков» между позвоночными артериями.
Иногда в полости основной артерии сохраняется на том или ином протяжении перегородка, представляющая собой остаток сомкнувшихся внутренних стенок позвоночных артерий. Чрезвычайно редко наблюдается также наличие анастомозов между отделами позвоночных артерий, находящимися в полости черепа. Обычно такого рода анастомозы располагаются у места слияния позвоночных артерий в основную, каудальнее они встречаются значительно реже. Лишь в 3 случаях. Гохштеттер и Шмайдель (1932, 1936) наблюдали их в области задних отделов олив продолговатого мозга. К крайне редким явлениям должно быть отнесено также наличие анастомозов между передними концами позвоночных артерий (Гохштеттер, 1937). Среди большого количества просмотренных Гохштеттером препаратов мозга взрослых ни в одном из случаев он не обнаружил анастомозов между головными концами позвоночных артерий (в области развилка основной артерии), но отмечал их у плодов.
Обычными вариациями основной артерии являются изменения в ее длине, зависящие от того, на кате*! протяжении слились между собой позвоночные артерии. Большая или меньшая величина осисшой артерии в свою очередь обусловливает и вариации в обхождении ее ветвей, особенно нижних мозжечковых артерий.
В большинстве случаев основная артерия разделяется у верхней границы варолиева моста на две, уже описанные выше задние мозговые артерии. Помимо их, основная артерия дает ветви, питающие средний: мозг, варолиев моет и мозжечок.
Непосредственно от верхнего конца основной артерии (кзади от места отхождения задней мозговой артерии) отходит верхняя мозжечковая артерия, место возникновения и путь которой, согласно описанию многочисленных 'авторов, довольно постоянны.
Исследования места обхождения этой артерии показали, что у человека, обезьян, собак, кошек и кроликов верхняя мозжечковая артерия в подавляющем большинстве случаев отходит одним стволом и в дальнейшем своем ходе распадается на две ветви (И. М. Григоровский, 1930; Б. К. Гиндце, 1947, и др.).
|
По данным А. Чернышева и И. М. Григоровского, а также А. Я. Подгорной (1936), С. С. Брюсовой (1938) и др., для человека характерно деление основного ствола верхней мозжечковой артерии на три и даже четыре вторичные ветви, что не встречается у животных. У человека и кошки (И. М. Григоровский, 1930; А. Я. Подгорная, 1936; Б. К. Гиндце, 1947), а также у гориллы и орангутанга (Б. К. Гиндце) может иметь место отхождение верхней мозжечковой артерии двумя самостоятельными стволами на одной из сторон, в то время как на другой соответствующая артерия отходит одним стволом. Подобные соотношения И. М. Григоровский наблюдал в 24% исследованных им случаев, причем на долю правой стороны приходилось 29%, а на долю левой — 18% всех случаев (рис. 10).
К наиболее редким явлениям принадлежит отхождение верхних мозжечковых артерий двумя стволами с обеих сторон, а также отсутствие этой артерии на одной из сторон.
| Рис. 10. Вариации верхней мозжечковой артерии (по Григоровскому). |
Ветви верхней мозжечковой артерии у человека снабжают передний край варолиева моста, ножки мозга, красное ядро, черную субстанцию, заднее двухолмие, ядро глазодвигательного нерва, спино-таламический тракт; распределяются они также по верхней поверхности червя и мозжечка. Верхняя мозжечковая артерия является главной артерией, снабжающей зубчатое ядро мозжечка.
Области распространения отдельных ветвей верхней мозжечковой артерии сильно вариируют. Разветвления ее широко анастомозируют с ветвями задней нижней мозжечковой артерии, часто снабжая кровью при недоразвитии последней соответствующие области. Верхняя мозжечковая артерия связана большим количеством анастомозов также с зад-не-передней мозжечковой артерией (рис. 11).
Вторая крупная ветвь основной артерии — н и ж н я я передняя м о з ж е ч к о в а я артерия — у человека в большинстве случаев отходит от основной артерии одним стволом, в дальнейшем разделяющими на две или три ветви. Однако отмечены случаи отхожденйя передави нижней мозжечковой артерии у человека, а также у собак двумя и даже тремя стволами. В последнем случае трений ствол является ветвью позвоночной артерии и в дальнейшем своем ходе все три ветви объединя-
ются в один общий ствол (Б. К. Гиндце, И. М. Григоровский) человеку, у обезьян описываемая артерия отходит чаще всего
|
одним стволом, чрезвычайно вариируя по месту отхождения, количеству и величине периферических ветвей.
У кошек эта артерия образует один общий ствол с нижней задней мозжечковой артерией [И. М. Григоровский, 1930, и др.]. Подобные же соотношения некоторые авторы, в. том числе и А. Я. Подгорная (1936), наблюдали у людей. Наконец, по данным А. Чернышева, у кроликов этой артерии установить не удается. Области распространения разветвлений передней нижней мозжечковой артерии вариируют по своей протяженности, охватывая задние и боковые отделы варолиева моста, боковые части полушарий мозжечка, где они широко анастомозируют с ветвями верхней и нижней задней мозжечковых артерий. Место отхождения третьей большой артерии, снабжающей мозжечок— задней нижней мозжечковой артерии, в различных случаях может быть отнесено и к основной, и к позвоночным артериям.
| Рис. 11. Артериальное кровоснабжение мозжечка и анастомозирование ветвей мозжечковых артерий в мягкой мозговой оболочке, а — базальная сторона: 1 — задняя соединительная артерия; 2 — верхняя мозжечковая артерия; 3 — задняя мозговая артерия; 4 — основная артерия; 5 — средняя . мозжечковая артерия; 6 — передняя спинальная артерия; 7 — нижняя мозжечковая артерия; 8 — позвоночная артерия; 9 — задняя спинальная артерия. б — вид сбоку: 1 — верхняя мозжечко-вая артерия; 2 задняя ветвь задней мозговой артерии; 3—задняя мозговая артерия; 4 — основная артерия; 5 — средняя мозжечйовая артерия; 6 — нижняя мозжечковая артерия; 7 - позвоночная артерия. |
Согласно описаниям большинства исследователей, задняя нижняя мозжечковая артерия в нормальных условиях возникает от верхней части по-зво'но'чной артерии [Лушка (Luschka), 1867; С. М. Огнева, 1944, и др.]. Это так называемое «низкое» отхож-дение артерии было отмечено в. 68% всех случаев, приведенных в большой работе И. М. Григоровского (1930).
В меньшем количестве случаев, но нередко, эта артерия отходит от основной артерии [С. М. Огнева, Генле (Henle), 1868]. Подобный же способ возникновения был обнаружен Шарли у некоторых животных (овец, лошадей и т. д.).
Что касается способа отхождения описываемой артерии у животных, то
в этом отношении данные А. Чернышева указывают на возникновение задней нижней мозжечковой артерии у обезьян и кроликов одним стволом от основной артерии. Для собак характерно отхождение артерии двумя самостоятельными стволиками, причем один из них берет начало от позвоночной артерии, другой — от основной.
В отдельных случаях А. Чернышев, А. Я. Подгорная и др. отмечали у человека происхождение нижней задней мозжечковой артерии одно временно от позвоночной и основной артерий. Стволы в дальнейшем или объединялись в, одну общую артерию, или шли совершению самостоятельно, параллельно друг другу.
|
Среди многообразных способов возникновения и хода этой артерии нужно отметить также нередкие случаи отсутствия ее с той или другой стороны (Блэкбурн, 1907; Б. К. Гиндце, М. Григоровский и др.). К особен-ностям этой артерии необходимо от-нести ее петлеобразное ветвление при прохождении, в большой мозжечковой цистерне. Обычно она делает одну или полторы петли, прежде чем перейти на поверхность мозжечка (рис. 12).
Главный ствол задней нижней мозжеч-
ковой артерии в дальнейшем, чаще всего у
человека, обезьяны, собаки, разделяется на
две, три и даже четыре вторичные ветви,
которые снабжают нижнюю поверхность
моззжечка и червя, сосудистое сплетение чет-
| Рис. 12. Вариации нижней задней мозжечковой артерии (по- Григоровскому). а — схема трехосновных типов, деления нижней мозжечковой артерии; б — схемы подгрупп первого основного типа ветвления нижней мозжечковой арте-рии на две вторичные ветви и на третичные ветви. |
вертoгo желудочка, а также может служить
источником кровоснабжения корешков
VIII нерва.
Основным источником кровоснабжения зубчато-го ядра мозжечка, по данным ряда исследователей, принято считать верхнюю мозжечковую артерию. Указывают также и на возможность питания этого образования из нижней передней мозжечковой арте-рни. В работе сотрудницы лаборатории М. Г. Привеса, А. В. Дроздовой в последнее время (1948) получены результаты, позволяющие ей зорить об участии в кровоснабжении зубчатого ядра всех мозжечковых артерий. А.В. Дроздова обнаружила вхождение верхней мозжечковой артерии в зубчатое ядро, в то время как обе другие мозжечковые артерии оплетают это ядро с поверхности и ветви их в глубине вещества ядра анастомозируют с ветвями верхней мозжечковой артерии-Данные, полученные при нейрохирургических операциях, пока не дают основания предполагать какой-либо другой источник кровоснабжения зубчатого ядра, за исключением верхней мозжечковой артерии.
Интересующихся кровоснабжением мозжечка мы отсылаем к работам упомянутых выше авторов, а особенно к работе С. С. Брюсовой (1940), в которой подробно изложена анатомия системы мозжечковых артерий, даны сведения о вариациях и описаны; зоны васкуляризации каждой из них. Хирурги найдут необходимые им подробности также в монографии Б. Г. Егорова (1949).
Основная артерия является главным источником, откуда берут на-
чало сосуды, снабжающие варолиев мост.
Вся совокупность артерий моста, может быть разбита на три группы. В первую группу входят с р е д и н н ы е артерии, ко второй и третьей соответственно относятся круговые короткие и круговые д л и н н ы е а р т е р и и.
Срединные артерии в количестве 4—6 отходят непосредственно от задней поверхности основной артерии и вступают в вещество варолиева места в радиальном направлении. Некоторые сосуды в своем распространении достигают дна ромбовидной ямки. Срединные артерии снабжа-ютзаднюю часть ядра глазодвигательного нерва, задний продольный пучок, текто-спинальный тракт, ядра блокового и отводящего нервов среднюю часть медиальной петли, поперечные мостовые волокна, пира-
мидные пути и медиальную часть трапецивидного тела.
Боковая поверхность основной артерии служит местом отхождения 4—5 круговых коротких и 2 круговых длинных артерий. Обе группы сосудов своими ветвями охватывают всю наружную поверхность варо-лиева моста. Отходящие от них артерии нагружаются в вещество
Рис. 13. Зоны снабжения продолговатого мозга отдельными артериями в верхних и нижних его этажах (по Фуа).
варолиева моста в радиальном направлении и снабжают средние ножки мозжечка, латеральную часть трапециевидного тела, верхнюю
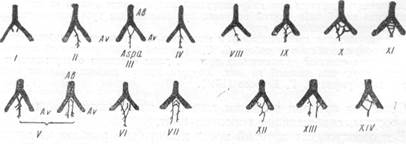
Рис. 14. Вариации передней спинальной артерии (по Стопфорду).
оливу, латеральную часть медиальной петли, ядро лицевого нерва, ядра VIII нерва, ядра тройничного нерва. От средней части основной артерии обычно отходит также a. auditiva interna (a. labyrinthica). Эта значительная по своей протяженности артерия проходит по вентральной и частью по боковой поверхности варолиева моста и вместе с комплексом VII, VIII черепномозговых нервов., а также n. intermedius входит во внутреннее слуховое отверстие. Артерия снабжает лабиринт и нижнесреднюю часть мозжечка.
Кровоснабжение продолговатого мозга обеспечивается передней спинальной артерией, ветвями, идущими от позвоночных артерий, от нижнего отдела основной артерии и частью от нижней задней мозжечковой артерии. Среди указанных артерий, описанных в соответствующих разделах данной главы, следует сказать несколько снов лишь о передней спинальной артерии (рис. 13).
Как видно из рис. 14, передняя спинальная артерия, представляющая собой крупную ветвь позвоночных артерий, имеет весьма разнообразные способы образования. В подавляющем большинствеслучаев она берет начало от каждой из позвоночных артерий и продолжается или в
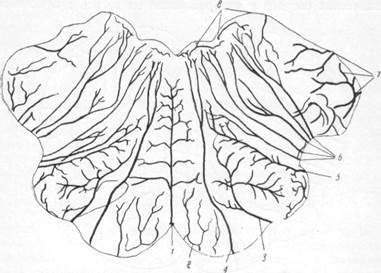
Рис. 15. Схема распределения крупных артериальных сосудов внутри продолговатого мозга на уровне верхней трети оливы
(по Бонне).
1, 2, 4 —срединные ветви; 3, 5 —артерии, снабжающие оливу; 6—ветви a. sulcus latera'is bulbi; 7 — артерии, снабжающие веревчатое тело; 8 — ветви задней спинальной артерии.
виде двух самостоятельных стволов, или образуетнепарный сосуд, сливаясь на уровне пирамид. Многочисленные варианты происхождения н строения этой артерии описаны рядом авторов (С. М. Огнева, Бонне и др.).
Тип кровоснабжения продолговатого мозга сходен с описанным выше для варолиева моста, т. е. продолговатый мозг получает кровь по срединным и круговым артериям, хотя ход этих сосудов менее правилен, чем в варолиевом мосту (рис. 15 и 16).
Срединные артерии возникают от задней поверхности основной, позвоночных и передней спинальной артерий и вступают в толщу мозгового вещества продолговатого мозга в виде внутримозговых радиальных сосудов, причем некоторые из них достигают дна четвертого желудочка. Эти артерии снабжают кровью пирамидный путь, медиальную петлю, предорзальный и задний продольный пучки, а также область расположения ядра подъязычного нерва.
Среди коротких круговых артерий продолговатого мозга можно выделить артерию латеральной ямки (a. fossae lateralis bulbi), которая отходит от основной артерии, идет вдоль нижней оливы в борозде,
располагающейся над этим образованием. Ветви этой артерии в количестве 4 или 5 входят в вещество продолговатого мозга и вместе с ветвями от нижней мозжечковой артерии распределяются в задне-боковых. отделах продолговатого мозга. Ветвями этих артерий снабжаются олива, желатинозная субстанция, нисходящий корешок тройничного нерва, ядра языкоглоточного и блуждающего нервов и вестибулярные ядра (рис. 15, 6). Часть сосудистых веточек проникает и мозговое вещество продолговатого мозга по ходу нервных волокон корешков черепно-мозговых нервов.
Изложенные выше данные о кровоснабжении продолговатого мозга. и варолиева моста дают описание общего характера вхождения крупных макроскопических сосудов и распределения их ветвей в этих областях
Рис. 16. Схема распределения крупных артериальных
сосудов внутри продолговатого мозга на уровне нижней
трети оливы (по Бонне). Анастомозы между артериями
так же как на рис. 15 не показаны.
В своей монографии, посвященной кровоснабжению продолговатого мозга, М. А. Захарченко (1911) писал: «Все изложенное — только общие контуры и порою только обрывки контуров той сложной и пестрой картины, детальная отделка которой еще долгое время будет служить. богатой темой для анатомического исследования».
Несмотря на то, что быстрое развитие нейрохирургиив последнее время требует обширных и детальных сведений о кровоснабжении всех отделов головного мозга, успехи, сделанные наукой в этом направлении, еще так незначительны, что слова, сказанные когда-то М. А. Захарченко, сохраняют свое значение до наших дней. В настоящее время мы можем сказать, что слова эти справедливы не только в отношении продолговатого мозга, но и в отношений варолиева моста, среднего мозга, подкорковых образований больших полушарий и т. д. Обзор имеющихся в литературе работ, освещающих вопросы кровоснабжения мозга, показывает, что основной интерес большинства исследователей
1 Подробности кровоснабжения варолиева моста и продолговатого мозга сосудами микроскопического порядка имеются в соответствующем разделе диссертационной работы С. М. Огневой (1944).
сосредоточивается до сих пор на анатомии и физиологии кровообращения в полушариях головного мозга и значительно менее на других отделах его.
Имеющиеся работы, посвященные этим отделам, ограничиваются описанием основных артериальных стволов, снабжающих то или другое образование. Совершенно не изучена сосудистая сеть мягкой мозговой оболочки почти всех этих отделов мозга (некоторым исключением является только мозжечок). Нет работ, касающихся вопросов тонкой сосудистой архитектоники в мозговом веществе, не начато еще изучение онтогенетического развития артериальной сети в мягкой мозговой оболочке и внутримозговой субстанции всех отделов за исключением полушарий головного мозга.
Разрешение этих вопросов является очередной задачей предстоящих исследований.
2. Анатомия вен мозга и венозных синусов
В то время как по вопросу о характере снабжения головного мозга артериальной кровью, привлекавшему внимание многочисленных исследователей вплоть до настоящего времени, накоплен значительный материал, сведения относительно венозной системы мозга в целом более скудны. Если изучению различных сторон проблемы артериального крово-бращения в мозгу различных животных и человека посвящено большое количество анатомо-гистологических и физиологических работ, то относительно венозной системы существует сравнительно незначительное количество работ, в основном относящихся к вопросам анатомии вен и венозных синусов мозга человека.
В литературе совершенно не затронут вопрос о строении венозной
части сосудистой сети мягкой мозговой оболочки, нет никаких данных
развитии венозной сети мозга и о соотношениях ее с артериальной
сетью мягкой мозговой оболочки и мозгового вещества. За исключением
исследований Пфайфера, которые, как мы увидим дальше, только
благодаря ошибочной классификации артерий и вен мозга оказались
посвященными изучению строения венозной сети мозгового вещества,
и работы Шлезингера (1939), не существует никаких специальных
гистологических работ по этому вопросу. Уже после того, как настоящая
монография была приготовлена к печати, сотрудница нашей лаборатории
Е. В. Капустина направила для опубликования данные своих работ по
указанным вопросам1. Внимание физиологов также сосредоточивалось
лишь на артериях мягкой мозговой оболочки, тогда как о венах в их
работах почти совсем не упоминается.
В учебниках и монографиях обычно весьма кратко и в самой общей форме сообщаются анатомические данные о венозной системе мозга человека без попытки дать какое-либо физиологической толкование особенностям венозного кровообращения в мозгу.
Отсутствие литературных данных поэтому вынуждает нас в настоящем разделе нашей работы ограничиться лишь общими сведениями по анатомии вен мозга человека.
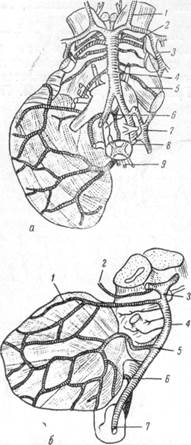
Вены мозга принято делить на систему поверхностных и систему глубоких вен. Считается, что поверхностные вены отводят кровь, еодер-жащую продукты обмена веществ серого и белого вещества полу-шарий головного мозга, а по глубоким венам оттекает кровь от подкорковых образований.
1 См. названия этих работ в литературном указателе.
Анатомо-гистологические данные показывают, что деление вен мозга, согласно этому принципу, является грубо схематическим. Поверхностные вены распределяются не только в коре и подлежащем белом веществе, но проникают далеко вглубь последнего вплоть до желудочков, где широко анастомозируют с ветвями глубоких вен. Распространение глубоких вен не ограничивается только областями подкорковых образований; они выходят за пределы этих образований в белое вещество извилин, где посредством многочисленных анастомозов соединяются с ветвями поверхностных вен. Наличие большого количества анастомозов между венами в мягкой мозговой оболочке и внутри мозгового вещества обеспечивает целостность в пределах системы поверхностных и глубоких вен и создает единство венозного кровообращения всего мозга в целом.
Трудно поэтому произвести строгую диференцировку венозного оттока в том или ином направлении, вследствие чего классификация вен по принципу распределения ветвей той или иной группы вен по областям утрачивает свое значение. В обычных условиях можно говорить лишь о преимущественном сосредоточении венозной крови ив той или иной области мозга, в какой-либо отдельной вене или группе их.
Несомненно, что сложная система анатомических путей оттока, служащих для выведения продуктов обмена веществ, существует в мозгу для обеспечения постоянства среды, окружающей нервную клетку.
Исходя из этого, естественно полагать, что повышение функциональной деятельности отдельных групп нервных клеток должно сопровождаться более быстрым отведением продуктов обмена веществ этих клеток, чем это имеет место в отсутствие повышения жизнедеятельности. При таких обстоятельствах для оттока крови будут вовлекаться не только венозные сосуды, выводящие продукты обмена обычно в определенном направлении, например, на поверхность мозга, но и другие пути, по которым венозная кровь начнет оттекать уже в ином направлении, например, в глубокие вены мозга.
Внутримозговые или радиальные вены, которые в обычных условиях отводят продукты обмена веществ из всей толщи серого и белого вещества мозга, поднимаются на его поверхность и в мягкой мозговой оболочке формируют густую сеть сосудов разнообразного калибра. Венозная сеть располагается в мягкой мозговой оболочке независимо от артериальной сети, направление венозных стволов ее не совпадает с направлением артерий в соответствующих областях. Более того, как будет видно из дальнейшего, артерии и вены в мягкой мозговой оболочке часто идут в совершению противоположных направлениях и не имеют того параллельного друг другу хода, как это наблюдается в мозговом веществе.
Образующиеся от слияния мелких вен крупные венозные стволы на поверхности мозга авторы классифицируют по группам, согласно самым разнообразным принципам.
Одни делят вены на передние, средние или задние, в зависимости оттех отделав полушарий головного мозга, где они проходят (Люшка, Крувилье). Другие предлагают классифицировать вены, расположенные в мягкой мозговой оболочке, исходя из направления их хода, и соответственно этому различают восходящие и нисходящие вены. При такой классификации вены основания выделяются в особую группу (Раубер, Тестю). Существует также распределение вен по группам в зависимости от тех синусов или пазух твердой мозговой оболочки, куда они впадают.
Мы в настоящего разделе нашей работы будем придерживаться
деления вен наружной поверхности мозга на группы соответственно направлению их хода, как это приведено у С. С. Брюсовой (1938).
С. С. Брюсова различает: 1) восходящие вены, к которым могут быть отнесены лобные вены, вены центральных извилин и теменно-затылочные вены; 2) систему средней мозговой вены; 3) группу височ-но-затылочных вен.
|
При рассмотрении вен, идущих в восходящем направлении и впадающих в верхний продольный синус, разные авторы отмечают различное количество крупных венозных стволов. По данным С. С. Брюсовой, число их колеблется от 5 до 11, причем наиболее, часто можно видеть 6—7 стволов. Сарджснт (Sargent, 1915) насчитывает всего 4 крупных вены, Шенкин различает 8—12 (Shen-kin, 1948). По мнению Бейли (Bailey, 193B), несмотря на большую изменчивость в отношении общего количества крупных вен на верхней наружной поверхности мозга (от 6 до 12), обычно наблюдается 4—5. Крувилье насчитывает 7—8 вен, Тестю - 8—12, Ге'нле — 10—12, Люшка — 12—15. Вены эти имеют калибр от 1,5 до 5 мм и разбиваются, как мы уже видели, на 3 группы.
| Рис. 17. Вены наружной поверхности мозга и характер впадения их в верхние продольный синус (по Бейли). 1 — лобные вены; 2 — верхний продольный синус; 3 — вена прероландовой борозды; 4— парасагиттальнме лакуны; 5 — вена роландовой борозды; 6 — затылочные вены. |
Среди группы поверхностных вен, идущих в восходящем направлении, различают:
I. Лобные вены (v. frontales;
рис. 17, 1), собирающие кровь с
верхней поверхности лобной доли
и направляющиеся вперед и вверх
к верхнему продольному синусу,
в который и впадают, частью не
сколько отклоняясь кзади. Диаметр их колеблется от 1 до 4 мм
(С. С. Брюсова).
II. Вены центральных извилин в количестве 2 или 3, ход которых
обычно совпадает с направлением соответствующей извилины (v. prae-
rolandica и v. rolandica; рис. 17, 3, 5). Более значительные по своему
калибру (от 2 до 5 мм), эти вены собирают кровь из бассейнов средней
и передней мозговых артерий. При выходе из мягкой мозговой оболочки
ка протяжении 2—3 см вены идут свободно в субарахноидальном про
странстве, изгибаясь кпереди, принимают косое направление и впадают
в нижний край верхнего продольного синуса. Среди группы центральных
вен в свою очередь различают:
1. Вену прецентральнойборозды (v. praecentralis, или v. praerolan-
dica), обеспечивающую отток венозной крови главным образом с перед
ней центральной извилины и с задних отделов верхней и средней лоб
ных извилин. Перед впадением в синус эта вена сливается с веной,
идущей с медиальной поверхности полушария.
2. Вену роландовой борозды (v. rolandica). Эта вена сосредоточи-
вает в себе кровь с заднего края передней центральной извилины, но служит также и для оттока части венозной крови с задней центральной извилины. Подобно предыдущей, вена роландовой борозды сливается с веной медиальной поверхности перед впадением в верхний продольный синус.
3. Вену постцентральной борозды (v. postcentralis), по которой продукты обмена веществ отводятся в основном с задней центральной
|
извилины, а также и от смежных с ней участков верхней и нижней теменных долек. Соответственно этой вене с медиальной поверхности подходит еще венозный ствол, вливающийся вместе с ней в синус.
| Рис. 18. Вены наружной и медиальной поверхности полушарий (по Бейли). а. 1 — вена Троларда; 2 — вены роландовой борозды; 3 — вена Лаббе; 4 — средняя мозговая вена; 5 — ана стомоз между ветвями лобных вен и ветвями средней мозговой- вены. б. 6 — вена Галена; 7 — затылочная вена; 8 — основная вена. |
III. Вены теменно-за-тылочной области (v. occi-pitales; рис. 17, 6) характеризуются, по данным С. С. Брюсовой, ветвистостью строения и формированием ствола из многих веток. Вены этой группы в количестве 1—2—3 стволов имеют диаметр от 2 до 4 мм, собирают кровь с теменных и затылочных извилин. Подобно предыдущим, описываемые вены при подходе к синусу за несколько сантиметров до него делают изгиб вперед и впадают в него под острым углом. Эта группа вен включает в себя: 1) переднюю теменную вену; 2) заднюю теменную вену; 3) затылочную вену.
Вены, отводящие
кровь в нисходящем направлении, изливают ее в поперечный синус (sinus transversus), верхний каменистый синус (sinus petrosus superior) и в вену Галена. Входящая в состав этой группы передняя височная вена обеспечиваетотток крови со средних отделов височных извилин. Продукты обмена веществ мозгового вещества задних отделов тех же извилин, а также угловой извилины и нижних затылочных извилин выводятся по задней височной вене. Венозная кровь от нижних затылочных извилин изливается также в нижнюю затылочную вену (v. occipitalis inf.), впадающую в вену Галена перед самым вхождением последней в прямой синус. С нижней поверхности
лобной доли вены направляются к нижнему продольному или к пещеристому синусу.
|
Между описанными группами вен, отводящими главную массу крови в восходящем или нисходящем направлении, располагается средняя мозговая вена (v. ccrebri media; рис. 18, 4, и рис. 19, 1, 9). По своему ходу она обычно совпадает с направлением сильвиевой борозды и широко анастомозирует с ветвями восходящих и нисходящих вен. Область распределения ветвей этой вены охватывает, идя спереди назад, края лобной, теменной и височных долей и внутреннюю поверхность височной доли - островок. Из всех этих областей венозная кровь отводится в верхний каменистый или пещеристый (sinus caverno-sus) синусы.
Поверхностные вены полушарий головного мозга большей частью имеют соответствующие им вены на медиальной поверхности полушарий; это с особой отчетливостью видно на мозгах эмбрионов.
При переходе выпуклой поверхности полушария в медиальную обе вены соприкасаются и в одних случаях впадают в синус раздельными стволами, в других— сливаются в одну вену, которая затем уже впадает в синус.
| Рис. 19. Вены основания мозга (по Бейли). 1 — средняя мозговая вена; 2 — основная вена; 3 — латеральные затылочные вены; 4 — внутренние затылочные вены; 5 — прямой синус; 6 — боковой синус; 7 — латеральные затылочные ьены; 8 — вена Лаббе; 9 — средняя мозговая вена. |
Примером может служить уже описанное слияние вены ро-ландовой борозды, а также постцентральной и других вен с соответствующими им венами, соби-рающими кровь с медиальной поверхности полушария. На рис. 18
можно видеть, что отток венозной крови с медиальной поверхности полушария происходит не только в верхний продольный синус, но и в противоположном направлении в основную вену (v. basilaris, рис. 18, 8 и рис. 19, 9). Основная, или, по другой терминологии, нижняя, мозговая вена собирает кровь, кроме того, с передних отделов поясной извилины, с клина и задних отделов поясной извилины и несет ее в вену Галена (v. cerebri magna; рис. 18, 6).
Характерной особенностью вен, располагающихся в мягкой мозговой оболочке на поверхности мозга, является большое разнообразие количества крупных венозных стволов, впадающих в те или иные синусы, величина просвета этих стволов, способ их хода и характер ветвления. Наиболее же отличительная черта поверхностных вен мозга заключается в наличии большого количества венозных анастомозов различного диаметра, соединяющих между собой ветви одной и той же вены и разветвления различных стволов.
Если в вопросе о строении артериальной сети мягкой мозговой оболочки до последнего времени оставалось много неясных мест, касавшихся главным образом размера и количества анастомозов, то в отношении-
вен мягкой мозговой оболочки эти вопросы могут считаться окончательно установленными и не могут вызывать каких-либо сомнений.
Начиная с XIX века (Вик д'Азир, 1876) и до наших дней, все исследователи указывают на наличие в мягкой мозговой оболочке подлинной сети венозных сосудов, охватывающей всю поверхность полушарий большого мозга, мозжечка и других отделов головного мозга. Это может считаться прочно установленным благодаря многочисленным наблюдениям анатомов и нейрохирургов, несмотря на то, что в литературе пока нет специальных работ, освещающих вопросы анатомического строения венозной сети мягкой мозговой оболочки.
Непрерывность в пределах этой сети осуществляется за счет большого количества анастомозов, достаточно крупного калибра для того, чтобы служить окольными путями в случае возможного перемещения крови в различных направлениях при нарушении нормальной ее циркуляции. Изучение анатомических путей, служащих для выведения продуктов обмена веществ из мозгового вещества, показывает, что венозная кровь из одних и тех же извилин мозга может оттекать по двум или даже трем различным направлениям. Так, например, в нормальных условиях от двух верхних третей центральных извилин кровь оттекает в верхний продольный синус, а от нижней трети — в среднюю мозговую вену. То же можно сказать и о лобных, затылочных и других отделах полушарий головного мозга. От угловой извилины кровь одновременно направляется в три различных венозных резервуара — в верхний продольный синус, в поперечный синус и в среднюю мозговую вену.
Существование многочисленных анастомозов может обеспечить в Широких пределах передвижение венозной крови при отсутствии возможностей оттока ее в каком-либоиз обычных направлений.
Помимо наличия большого количества анастомозов различного диаметра, одной из особенностей венозной сети мягкой мозговой оболочки является существование крупные анастомозов, по своим размерам не уступающих или лишь незначительно уступающих стволам основных вен. Таковы, например, так называемые вены Лаббе иТроларда (рис. 18, 3,1).
Вена Лаббе, служит для непосредственного оттока крови из средней мозговой вены в поперечный синус или в противоположном направлении. Вена Троларда представляет собой коммуникационный путь между средней мозговой веной и верхним продольным синусом через вену роландо-вой борозды. Оба эти крупных анастомоза имеют просвет, одинаковый на всем их протяжении, обнаруживая иногда расширения у места соединения со средней мозговой веной или синусом. Последнее обстоятельство, по мнению М. Б. Копылова (1948), является доказательством возможности перемещения крови в них в том и другом направлении.
Основная масса венозной крови от подкорковых образований собирается в систему вены Галена. На рис. 20 можно видеть, что передние и задние терминальные и поперечные вены (рис. 20, 3, 6, 5), собирающие-кровь с хвостатого ядра и с прозрачной перегородки (septum pellucidum; рис. 20, 4), пересекают поверхность хвостатого ядра и в области бокового желудочка меняют направление своего хода и впадают во внутреннюю вену мозга (v. cerebri interna; рис. 20, 2). Кзади от места их впадения в ту же вену изливается кровь из вены сосудистых сплетений.
Парные внутренние мозговые вены, объединяясь, образуют вену Галена (v. cerebri magna; рис. 21, 1). Помимо указанных вен, вена Галена сосредоточивает в себе венозную кровь, поступающую по венам-зрительного бугра, аммонова рога, венам белого вещества, располагающимся по сторонам мозолистого тела.
По пути вены Галена к прямому синусу она принимает в себя также вены мозолистого тела (v. post, corporis callosi), верхнюю среднюю вену мозжечка (v. cerebelli superior mediana), основную вену и внутреннюю, затылочную вену.
Мы уже видели, что глубокие вены мозгового вещества вступают в
соединение с ветвями радиальных вен, выводящими продукты обмена
веществ серого и белого вещества мозга. Анастомозы между ветвями той
и другой системы, располагающиеся в белом веществе, могут служить.
непрямыми путями для оттока венозной крови в том или ином направлс-
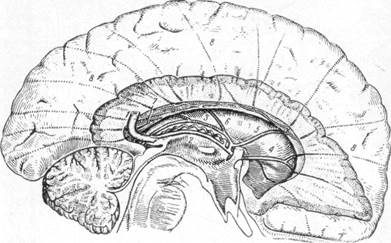
Рис. 20. Схема оттока венозной крови с подкорковых образований
(по Шлезингеру).
1 — вена Галена; 2 — внутренняя вена мозга; 3 — задняя терминальная
вена; 4 — вена прозрачной перегородки; 5 — передняя терминальная вена;
6 — поперечная вена хвостатого тела; 7 — продольные вены хвостатого
тела; 8 — внутримозговые анастомотические вены.
л в случае затруднения ее движения по обычным венам. Но, как показывают патологоанатомические исследования, размер анастомозов в белом веществе не настолько значителен, чтобы они могли играть существенную роль в перемещении крови от подкорковых образований к поверхности мозга или обратно.
В обычных условиях отток венозной крови из коры и белого вещества происходит в поверхностную сеть венозных сосудов мягкой мозго-вой оболочки, а продукты обмена веществ из подкорковых образований преимущественно выводятся системой вены Галена.
Однако, согласно анатомо-гистологическим данным некоторых авто-ров (см., например, Шлезингер, 1939), глубокие вены мозга сообщаются с крупными венозными стволами на поверхности мозга или с венозными пазухами посредством многочисленных каналов, проходящих через всю толщу белого и серого вещества. Эти каналы представляют собой сосуды диаметром до 0,5 мм, незначительно изменяющиеся на всем протяже-нии их хода от коры до околожелудочкового белого вещества (рис. 20, 8, и рис. 21, 5). Характерной особенностью описываемых коммуникацион-
ных путей является тот факт, что эти вены почти не ветвятся по всему своему ходу и лишь изредка вступают между собой в соединение посредством небольших венул, отходящих от них под прямыми углами (рис. 22, 10).
Вены такого рода, следовательно, являются приспособлением, обеспечивающим более широкие возможности для перемещения крови з обоих направлениях. Излагаемые дальше экспериментальные данные,
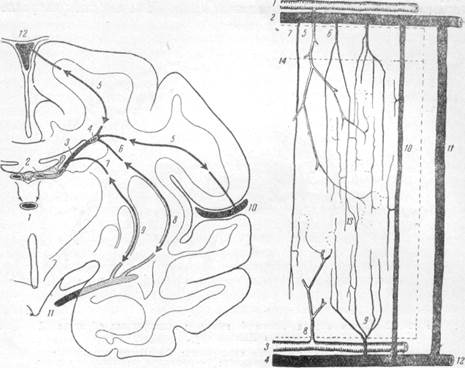
|
Рис. 21. Схема анастомозов, соединяющих
систему вены Галена с венами наружной
поверхности мозга (по Шлезингеру).
1 — вена Галена; 2 — внутренние мозговые вены; 3 — поперечная вена хвостатого тела; 4 — продольная вена хвостатого тела; 5 — вены, соединяющие вену Галена с верхним продольным синусом и поверхностными ве-
нами сильвиевой борозды; 6—7 — верхние наружные и внутренние вены чечевице-обрасного ядра; 8 и 9 — нижние наружные и внутренние вены того же ядра; 10 — поверхностные вены сильвиевой борозды; 11—
глубокие вены той же борозды; 12 — верхний продольный синус.
Рис 22. Схема, иллюстрирующая богатство анастомотических связей в венозной системе мозга (по Шлезингеру).
I — артерия мягкой мозговой оболоч
ки; 2 — вена мягкой мозговой оболоч
ки; 3 — артерия подкорковых узлов;
4 — ветвь вены Галена; 5 — артерия
коры; 6 — вена коры; 7 — кортикаль
ная и длинная субкортикальная вена;
8 - субвентршсулярная артерия; 9 —
субвентрикулярная вена; 10 — интра-
церебральные анастомотические вены:
II — экстрацеребральные анастомоти
ческие вены; 12 - вена Галена; 13 —
анастомозы между венами; 14 — гра
ница коры и белого вещества.
полученные в нашей лаборатории, позволяют считать эти вены следствием закрепления в венозной системе взрослого структур, характерных для его эмбрионального состояния. Подобного рода длинные сосуды, пронизывающие всю толщу стенки мозгового пузыря вплоть до матрикса и на некотором расстоянии соединяющиеся между собой небольшими по калибру поперечными сосудами, являются характерными для определенных ранних стадий развития сосудистой системы.
Все вышеизложенные данные позволяют заключить, что в венозной
системе наблюдаются различного рода анастомозы. Густая венозная сеть, располагающаяся в мягкой мозговой оболочке, включает в свой состав большое количество анастомозов различного диаметра, причем диаметр анастомозов может быть значительным. В то же время в коре и белом веществе крупные анастомозы встречаются сравнительно редко и взаимодействие сетей серого и белого вещества, а также венозной сети последнего с системой глубоких вен осуществляется главным образом посредством капиллярного русла и анастомозов прекапиллярного размера.
В особую группу должны быть выделены анастомозы, являющиеся значительными по калибру стволами, непосредственно соединяющими между собой в мозговом веществе глубокие и поверхностные вены, а на поверхности мозга - отдельные венозные синусы. Отсутствие физиологических данных, на основании которых можно было бы составить представление о путях движения венозной крови в обычных условиях и при патологических состояниях мозга, не дает возможности притти к определенному мнению о физиологическом значении всех отмеченных нами коммуникационных анастомозов. Можно лишь предположить, что наличие их в венозной сети мозга вызвано необходимостью существования одного и того же давления в системе поверхностных и глубоких вен мозга, а также и в системе венозных пазух твердой мозговой оболочки. Изменение давления в той или другой системе вызывает перемещение крови по крупным анастомозам и таким образом вновь создает состояние равновесия во всей венозной сети мозга в целом.
Вены при переходе из мягкой мозговой оболочки в синус свободно располагаются в субарахноидальном пространстве, причем в лобных отделах свободная часть вены может достигать 4 см, в задних отделах мозга она обычно не превышает 1 см. Иногда вены собираются под нижней поверхностью лакун продольного синуса, никогда в них не впадая.
В нейрохирургической практике встречаются случаи, когда отмечается впадение части вен наружной поверхности мозга не в синус, а в твердую мозговую оболочку, не доходя 3—4 см до синуса. В таких случаях зены идут к синусу на протяжении 3—4 см в нижнем листке твердой мозговой оболочки.
Среди работ, относящихся к венозной системе мозга, лишь одна работа К. Д. Балясова (1927) специально посвящена детальному описанию анатомии вен мозжечка.
Использовав данные, полученные при инъекции застывающей массы в вены мозжечка взрослых и детей, К. Д. Балясов показал, что вены мозжечка, подобно венам полушарий головного мозга, характеризуются большим разнообразием в отношении их количества, размера и положения.
О широких пределах возможных вариаций с достаточной убедительностью свидетельствует тот факт, что общее количество поверхност-ных вен мозжечка колеблется от 6 до 22. Однако при таких значительных размерах колебаний все же можно выделить постоянно встречающиеся вены и вены, встречающиеся далеко не на всех препаратах, — вариирующие.
Что касается размеров вен поверхности мозжечка, то и здесь па-блюдается большое разнообразие. Как удалось заметить, вены, впадающие в краевой синус, имеют меньший диаметр, чем вены, изливающие кровь в синусный сток (confluens sinuum).
В расположении вен какой-либо видимой закономерности установить также невозможно. Несмотря на то, что вены распределяются по
мозжечку без особого порядка, все они объединяются между собой большим количеством анастомозов, благодаря чему поверхность мозжечка, так же как и поверхность полушарий большого мозга, оказывается покрытой сплошной сетью венозных сосудов. Помимо подлинного анастомозирования между собой (сосудами крупного диаметра), поверхностные вены мозжечка анастомозируют с венами спинного и продолговатого мозга, с венозной сетью варолиева моста и четверохолмия, а также с венами большого мозга.
Вены, собирающие кровь с верхней поверхности мозжечка, с боковой поверхности ножек мозга и четверохолмия, с варолиева моста, а так-
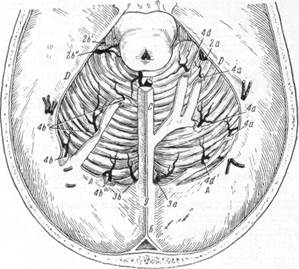
Рнс. 23. Схема оттока венозной крови с верхней поверхности мозжечка (по Балясову).
А — поперечный синус; Б — сагиттальный синус; С — прямой синус; D — каменистый синус; G — венозный сток; О — вена Галена; 2а и Зb', 2b"—клочковые вены; За и Зb — вены, впадающие в сток; 4а, 4b и 4Ь"—вены мозжечкового намета.
же и с нижней поверхности мозжечка объединяются в так называемые клочковые вены, впадающие в верхний каменистый синус (рис. 23, 2а, 2b', 2b"}.
С верхней поверхности мозжечка кровь собирается также парной или непарной венами, вливающимися в задний участок поперечного синуса (рис. 23, 4b").
В венозный сток впадают 1—2 вены, собирающие кровь с нижней поверхности мозжечка. Обычно по ходу этих вен к ним присоединяется веточка, несущая кровь с верхней поверхности (рис. 23, За, Зb).
Венозная кровь с квадратной дольки мозжечка вместе с кровью от миндалины собирается в 1—4 коротких венозных стволика, впадающих в краевой синус дорзально и латерально от продолговатого мозга (рис. 24, 7). С двубрюшной дольки и клочка венозная кровь по непарной или парным венам впадает в нижний каменистый синус (рис. 24, 2а, 2b). Наконец, сигмовидный синус принимает в себя венозную кровь, с миндалин мозжечка, прилежащих частей продолговатого мозга и с квадратной доли.
Среди данных, полученных К. Д. Балясовым, обращает внимание отсутствие впадения мозжечковых вен в затылочный и поперечные синусы у взрослых, тогда как у младенцев указанные синусы принимают по одной, две и даже три вены.
Вены мозжечка не имеют определенного участка впадения в синус и могут вливаться в него в самых различных местах, причем впадение их может быть или непосредственным, или после предварительного прохождения в толще мозжечкового намета.
Венозные синусы или пазухи, располагающиеся в дупликатуре твердой мозговой оболочки, служат резервуарами, воспринимающими кровь, поступающую в них по мозговым венам.
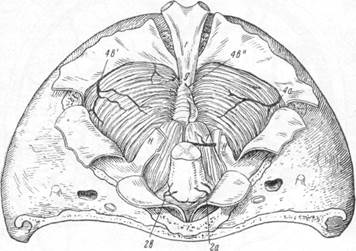
Рис. 24. Схема оттока венозной крови с нижней поверхности мозжечка (по Балясову).
Н — краевые синусы; G — сток; F — затылочный синус; 2а и
и 2в — вены, впадающие в нижний каменистый синус; 4b"—вена,
впадающая в прямой синус; 4а и 4b' — вены намета; 7 — вена
краепого синуса.
Представляя собой непосредственное продолжение вен мозга, венозные синусы в то же время резко отличаются от них по своему строению. В то время как мозговые вены имеют тонкую податливую стенку, благодаря чему просвет их может меняться под действием целого ряда факторов и в первую очередь вследствие изменения кровяного давления, стенки венозных синусов, состоящие из соединительной ткани с примесью эластических волокон, чрезвычайно плотны и мало податливы (Н. Н. Бурденко, 1927; М. Д. Злотников, 1947). Благодаря последнему обстоятельству на разрезах стенки синусов зияют.
Неподатливость стенок синусов, обеспечивающая свободное прохождение крови по ним, обусловливает особую роль их в механизме кровообращения внутри черепа. Вместе с тем неспадающиеся стенки представляют собой чрезвычайно неблагоприятный момент в случаях различного рода травм черепа, сопровождающихся нарушением целости сосудистой системы. Туго натянутые листки твердой мозговой оболочки, являющиеся стенками венозных синусов и образующие одно целое с впадающими в них венами, создают большие хирургические затруднения при попытках возместить дефект их стенок (Н. Н. Бурденко, 1927).
Подобно венам, синусы мозга обнаруживают большое разнообразие вариаций. Прежде всего это относится к величине просвета синуса. Известно, что для детей в первые годы жизни характерны более широкие синусы, чем это имеет место у взрослых. Нормальные свои размеры венозные синусы приобретают лишь с появлением и развитием диплоэ-тических вен (М. Б. Копылов, 1948).
В литературе описаны случаи, когда просвет синусов был весьма незначителен [А. Раубер, 1919; Ф. И. Валькер, 1922]. В отдельных,
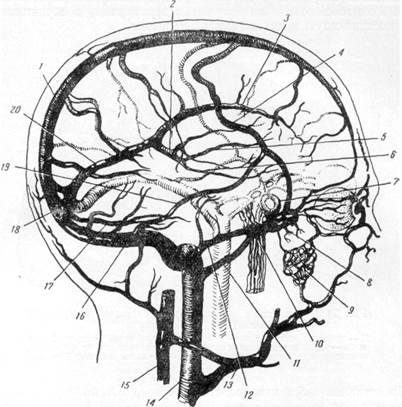
Рис. 25. Схема венозных синусов твердой мозговой оболочки
(ло Шенкину).
1 - верхний продольный синус; 2 — вена Галена; 3 — нижний продольный синус 4 — вены Троларда; 5 — внутренняя мозгоьая вена; 6 — основная вена; 7— интеркавернозный синус; 8 — пещеристый синус; 9 — крыловидное сплетение; 10 — основное сплетение; 11—нижний каменистый синус; 12—-верхний каменистый синус; 13 — общая лицевая вена; 14 — внутренняя яремная вена; 15— наружная яремная вена; 16 — правый поперечный синус; 17 — затылочный синус; 18—Torcular herophili; 19 — вена Лаббе;
20 — прямой синус.
правда, очень редких случаях наблюдается отсутствие верхнего продольного синуса. Вариации отмечаются также и в отношении расположения отдельных синусов. В подавляющем большинстве-случаев верхний продольный синус (sinus longitudinalis superior; рис. 25), являющийся наиболее мощной из венозных пазух, представляет собой непарное образование, располагающееся по средней линии в верхнем крас серповидного отростка. Верхний продольный синус идет от слепого отверстия (foramen coecum) дугообразно назад и у внутрен-
ней затылочной бугристости (protuberantia occipitalis intema) впадает в заднюю часть поперечного синуса (sinus transversus), отклоняясь при, этом вправо или влево.
Такое смещение верхнего продольного синуса в ту или другую сто-
рону имеет место приблизительно в 50% всех случаев. Наиболее часто местом впадения его является правый поперечный синус, причем к левому отходит небольшой канал. Наблюдается также деление синуса на
правую и левую ветви, продолжающиеся затем в соответствующие попе-речные синусы. В отдельных случаях можно отметить вторичное слияние правой и левой ветви синуса, в дальнейшем вновь распадающихся и сливающихся в поперечные синусы (Н. Н. Бурденко, 1927). В. Н. Шев-куненко описал редкий случай удвоения верхнего продольного синуса, причем каждая из пазух непосредственно продолжалась в поперечный^ синус своей стороны.
На поперечном разрезе верхний продольный синус имеет трехгран-ный просвет, постепенно увеличивающийся спереди назад. Особенностью уса является наличие лакун, представляющих собой расширение ду-пликатуры твердой мозговой оболочки, расположенных по краю синуса и соединяющихся с ней рядом небольших отверстий (рис. 17).
Лакуны отличаются большой вариабильностью как в отношении положения, так и в отношении величины. Иногда они настолько выражены, что ведут к значительному рассасыванию костей черепа, расположенных над ними. Помимо сообщения с продольным синусом, лакуны
соединяются с боков с менингеальными венами и с верхней стороны с диплоэтическими венами. Внутренняя нижняя стенка лакун бугриста-вследствие проникновения в нее пахионовых грануляций, представляющих собой разрастания паутинной оболочки.
Как уже указывалось, верхний продольный синус имеет связь со средней мозговой веной посредством вены Троларда. Второй анастомоз находится между его передним концом и венами полости носа.
Поперечные синусы, являющиеся на той или другой стороне продолжением описанного выше синуса, собирают почти всю венозную кровь черепной полости, благодаря тому, что в них непосредственно или через, другие синусы сливается кровь из всех остальных венозных синусов.
Оба поперечных синуса, имеющие на разрезе треугольную форму с основанием, сильно выпуклым наружу, проходят по заднему краю моз-
жечкового намета от внутренней затылочной бугристости до пирамидки
височной кости, где они загибаются вниз и носят название сигмовидных (sinus sigmoideus).
Сигмовидные синусы доходят до яремного отверстия (foramen jugu-lare) и, пройдя его, переходят во внутренние яремные вены (v. jugula-ris interna).
В большинстве случаев правый поперечный синус имеет просвет бо-лее широкий, чем соответствующий синус противоположной стороны, что, как мы уже видели, совпадает с более часто наблюдающимся слиянием верхнего продольного синуса с правым поперечным синусом. Место слия--верхнего продольного синуса с поперечным, располагающееся на внутренней затылочной бугристости, в 20—25% всех случаев (М. Д. Злот-ников, 1947) образует так называемый венозный сток (sinuum confluens). В 8% (по А. Рауберу, в 4 случаях из 50) в синусный сток вливается также прямой и затылочный синус.
Прямой синус (sinus fectus; рис. 25, 20) располагается по месту. соединения большого серповидного отростка (falx cerebri) с мозжечко-вым наметом. Благодаря тому что в начальный отдел его впадает вена
Галена (рис. 25, 2) и нижний продольный синус (рис. 25, 3), прямой синус по существу является продолжением нижнего продольного синуса.
Наблюдения говорят,
Дата добавления: 2015-05-16; просмотров: 1575;