Основные механизмы всасывания и транспорта веществ
Химус продвигается от двенадцатиперстной кишки вдоль всей тонкой кишки для полного переваривания и всасывания ворсинками и микроворсинками. Мышечная стенка тонкой кишки состоит из внутреннего циркулярного и наружного продольного слоев и совершает, как минимум, два типа сокращений: сегментацию и перистальтику. Сегментация вызывает перемешивание химуса, перемещая содержимое кишечника маятникообразно за счет периодических сокращений сегментов тонкой кишки. Перистальтика — это продвижение перевариваемого материала в направлении к толстой кишке. Данные мышечные сокращения контролируются нервной системой кишечника с модуляцией со стороны парасимпатической нервной системы и гормонов. Для лучшего переваривания и всасывания содержимое кишечника должно быть жидким, поскольку вода обеспечивает пространство для диффузии веществ из просвета кишки к поверхности энтероцита. Присутствующая в кишечнике вода поступает через рот с пищей, секретируется органами желудочно-кишечного тракта и тонкой кишкой (около 1.5 л). В тонкой кишке всасывается большая часть из 8.5 л воды, которые, в среднем, поступают в нее за сутки, и для окончательного всасывания до толстой кишки доходит около 0.5—2 л воды (рис.6-11).
Как только электролиты, пептиды, углеводы и липиды достигают энтероцитов, для их всасывания включаются различные механизмы. Это может быть даль
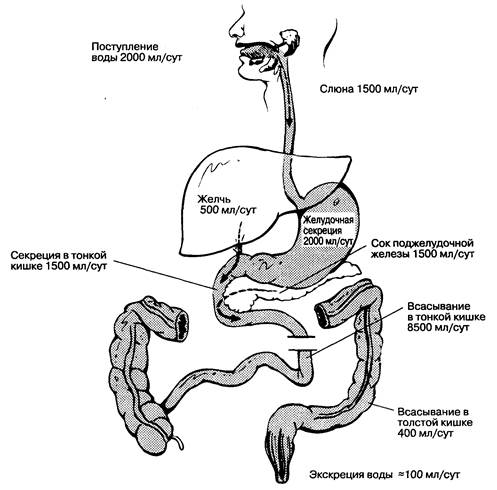
Рис. 6-11. Общий баланс воды в желудочно-кишечном тракте человека. (По: Berne R. M., Levy M. N., eds. Physiology. St. Louis: С. V. Mosby, 1983: 802.)
нейшее расщепление веществ ферментами энтероцитов, например дорасщепление углеводов и пептидов, а может быть непосредственное включение активного транспорта веществ из просвета кишки, например некоторых электролитов. Наружная мембрана энтероцитов обладает низкой проницаемостью для полярных химических структур, поэтому для переноса через нее питательных веществ необходимы специальные мембранные белки. Активный транспорт требует затрат энергии АТФ для переноса веществ против градиента их концентрации; например, против градиента концентрации работает Nа+,К+-АТФаза, локализованная в базолатеральном участке мембраны энтероцитов и перемещающая К+ в клетку, а Na+ из клетки. Белки в мембране клетки являются транспортными каналами, по которым осуществляется перемещение ионов в двух направлениях против градиента концентрации. Транспортные каналы могут находиться в "открытом" и "закрытом" состоянии. Примером их служит механизм перемещения ионов хлора на апикальном участке мембраны энтероцитов. Вторичный активный транспорт представляет собой комплекс активных и пассивных механизмов: он происходит за счет активного ионного (с затратой АТФ) транспорта, который формирует градиенты концентраций этих ионов и сочетается с пассивным транспортом энергетически "невыгодных" молекул, осуществляемым, например, транспортером Nа+/глюкоза, расположенным на апикальной поверхности энтероцитов. Na+,K+-АТФаза создает низкую концентрацию Na+ в клетке. Натрий стремится войти в клетку и с помощью транспортера Nа+/глюкоза входит вместе с глюкозой, которая, в отличие от натрия, идет против градиента концентрации (рис. 6-12). Этот транспорт происходит пассивно, но он невозможен без работы Nа+,К+-АТФазы. Na+/глюкоза-транспортёр является примером котранспорта (симпорт), поскольку и натрий, и глюкоза перемещаются в одном направлении. Обменный транспорт (антипорт) осуществляется с помощью белков, перемещающих однозарядные молекулы в разных направлениях. Поступление в энтероцит аминокислот, пептидов, витамина В12, желчных кислот происходит по принципу механизма котранспорта с Na+ как и описанный ранее транспорт глюкозы. Вода перемещается пассивно в направлении повышенного осмотического давления. После абсорбции воды оба сектора (просвет кишки и слизистая оболочка кишки) становятся изоосмотичными. Поскольку осмотическое давление, в основном, создается электролитами, перемещение воды регулируется преимущественно их транспортом. Главным ионом, управляющим перемещением воды, является Na+. Однако глюкоза и другие молекулы также осмотически активны, поэтому их абсорбция сопровождается абсорбцией воды.
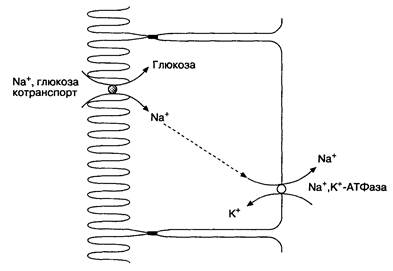
Рис.6-12. Абсорбция Na+ и глюкозы. На апикальном участке мембраны клетки находится Na+/глюкоза-транспортер, переносящий Na+ и глюкозу в клетку, после чего натрий выводится из клетки Na+,К+-АТФазой; глюкоза переносится через базолатеральный участок мембраны специальным переносчиком. Котранспортные механизмы с Na+ существуют для переноса аминокислот, ди- и трипептидов, некоторых витаминов группы В и желчных солей. (По: Yamada Т., Alpers D. П., Owyang С., Powell D. W., Silverstein F. E., eds. Textbook of Gastroenterology, 2nd ed. Philadelphia: J. B. Lippincott, 1995; 1:334.)
Дата добавления: 2015-02-23; просмотров: 1017;